



我们可以得出结论,一个有机体和它所经历的全部生物学相关过程,都需要建立在规模庞大的“多原子”结构上,从而防止“单原子”结构导致的偶然事件产生太大影响。“朴素物理学家”告诉我们,这一点非常重要,因为只有这样,有机体才能遵循足够精确的物理学定律,并在此基础上产生有规律和秩序的功能。从生物学的角度看,这些先验性的结论如何贴合到生物学的事实中呢?
乍看之下,人们倾向于认为这些结论无关紧要,比如30年前的一名生物学家也许就这样说过。尽管对于科普讲座来说,强调统计物理学在生物中和其他领域中的重要性别无二致是恰当的,但事实上这种观点不过是为人熟知的陈词滥调。
自然界中,任何高等生物的成年个体的躯体,以及构成躯体的单个细胞,都包含了“天文”数字数量的各种原子。我们观察到的每一种特定的生理过程,无论是发生在细胞内部的,还是细胞与细胞周围环境的相互作用,看上去——或30年前就看上去——也同样涉及海量的单原子和单原子过程,即使在刚才说明的 法则这种统计物理学的严格要求下,其巨大的数量也能保证相关的物理定律和物理化学定律的准确。
法则这种统计物理学的严格要求下,其巨大的数量也能保证相关的物理定律和物理化学定律的准确。
如今,我们知道这种观点是错误的。正如我们接下来会看到的,非常小的原子团,小到不完全符合统计学定律的原子团,在活的有机体内发生的极有秩序、有规律的事件中也起了主要作用。它们能够控制有机体在发育过程中可观察到的宏观特征,这些特征是有机体在发育过程中获得的,能够决定有机体发挥功能。在这些过程当中,生物定律都得到了清晰和严格的体现。
首先我必须要对生物学的情况进行总结,特别是遗传学的情况,换句话说,我要概括一个我不精通的学科的发展现状。我必须为自己内容的浅薄道歉,尤其是要向生物学家们道歉。但是,我也恳请你们暂时将主流观点放在一边。
生物学家已经积累了许多研究结果,一方面来自大量的、日积月累的繁育实验,这些实验之间完美地互相补充完善,具有史无前例的独创性;另一方面,来自通过精密的现代显微镜对活体细胞进行的直接观察。我们不能指望笨拙的理论物理学家能完成同样出色的研究。
我想用有机体的“模式”这个词来表示生物学家所说的“四维模式”的意义,它不仅是指有机体成年后或者任何特定阶段的结构和功能,还指有机体从受精卵到个体完全成熟开始繁殖的全部个体发育过程。现在,我们知道这个四维模式完全是由一个细胞即受精卵的物质结构决定的。而且,我们还知道本质上它完全是由这个细胞的一小部分结构也就是细胞核决定的。当细胞处于常规的“静止状态”时,细胞内的细胞核通常是网状染色质。但是在极为重要的细胞分裂(有丝分裂和无丝分裂,见下文)过程中,可以观察到细胞核会由一组被称为染色体的颗粒组成,通常呈纤维状或棒状,数量一般是8条、12条,人类是48条[1] 。不过我其实应该把染色体数目写成2×4,2×6,……,2×24,正确的说法也应该是有两组染色体,这样才是生物学家惯用的严谨表述。单个染色体有时候能够通过形状和大小清楚地被辨认和区分,但这里两组染色体几乎是一模一样的。我们如今已经了解到,其中一组来自母亲(卵细胞),另一组则来自父亲(精子)。
在显微镜下我们看到的染色体就像中轴骨一样,这些染色体中包含着某种代码文本,记录了个体未来发育和成年期身体功能的完整代码。每一组完整的染色体都含有全套的代码,所以一般来说,未来发展成个体的最初阶段的受精卵中包含两套完整的代码。
拉普拉斯设想过一种全知的智者,在他面前一切因果关系都能立即得出,我们把染色体纤维称为代码文本,是因为拉普拉斯设想的智者能够根据它的结构判断出在合适的条件下受精卵能够发育成黑公鸡还是花母鸡,能发育成苍蝇还是玉米、杜鹃花、甲虫、小鼠或是女人。我们可能要补充一点,卵细胞的外表看上去往往非常相似,即便它们略有不同,比如鸟类和爬行动物的卵就相对大很多(它们的卵如此巨大,不过是因为营养物质丰富),也根本比不上染色体结构的差异之大。
当然,代码文本这个词也过于狭隘了。染色体结构同时也是促进卵细胞未来发育的工具。它们既是规则的制定者也是执行者,换个比方,它们既是建筑师的蓝图也是建造者的工艺。
个体发育中染色体的行为是什么样的呢?
一个有机体的生长会受到细胞分裂的持续影响。这样的细胞分裂称为有丝分裂。在一个细胞的整个生命周期当中,有丝分裂出现的频率并不如人们想象的那样高,因为我们的身体是由数量庞大的细胞构成的。在生命初期,细胞数量的增长非常迅速。卵细胞会分裂成2个“子细胞”,然后会继续分裂成4个细胞,然后是8个、16个、32个、64个,以此类推。
在生长过程中,人体中各个部分的分裂频率并不会保持一致,所以会打破上面的增长规律。不过通过它们的增长规律,我们可以推断出,平均只需要50到60次连续分裂,就足以产生成年人类体内的细胞总量,或者如果把一生中细胞的更替也算进去,大概需要分裂出10倍于成年人体内细胞总量的细胞。因此,平均来说,我的任何一个体细胞都只是受精卵的第50代或60代“后代”细胞。
染色体在有丝分裂中的行为是什么样的呢?它们会进行复制,两组染色体、两套代码都会进行复制。人们利用显微镜对这个过程进行了大量研究,人们对于这个过程也非常感兴趣,但是它过于复杂,这里就不进行详细阐述了。这个过程的关键点是,两个子细胞都能够获得两组完整的、和母细胞完全一样的染色体。所以从染色体数量来讲,一个人所有的体细胞都是一模一样的。
尽管我们对这种机制了解得很少,但我们知道这与有机体的功能密切相关,每个细胞都有完整的一套(两份)代码文本,即便是不重要的细胞。
不久前,报纸上有这样一个报道:蒙哥马利将军在非洲的战役中,要求军队中每名士兵都要知晓其全部作战方案。如果报道内容完全真实(这种可能性相当大,因为他的士兵都是高智商的可靠之士),就为我们讨论的内容提供了一个绝佳的类比,每名士兵相当于一个细胞。最令人吃惊的是,有丝分裂过程中,细胞始终保持着两套染色体。遗传机制的显著特征是通过唯一一种与这条规则不符的细胞分裂凸现出来的,我们接下来要讨论这个话题。
个体发育完成之后,有一群细胞会被保存下来,以便在个体达到性成熟时产生配子用于个体繁殖。
根据个体性别不同,产生的配子分别是精细胞和卵细胞。所谓“保存”指的是,这些细胞不会有任何其他功能,同时进行的有丝分裂次数也更少。这种独特的细胞分裂叫做减数分裂,当生物个体性成熟后,一般在有性生殖发生前的很短一段时间内,会通过这种分裂方式让保留下来的细胞产生配子。减数分裂过程中,母细胞中的两个染色体组只是分成两个单独的染色体组,分别进入两个子细胞即配子当中。换句话说,有丝分裂过程中染色体翻倍的现象并不会出现在减数分裂当中,染色体数目保持恒定,因此所有配子都只有一半数量的染色体,只有一套完整的代码,而不是两套代码,比如人类的配子当中只有24条染色体,而不是2×24也就是48条。
只有一个染色体组的细胞叫做单倍体(haploid,来源于希腊语,意思是单数)。所以配子都是单倍体,而体细胞为双倍体(diploid,来源于希腊语,意思是双数)。体细胞当中包含三个、四个或概括地讲包含多个染色体组的个体偶尔也会出现,它们被称为三倍体、四倍体,以及多倍体。
有性生殖过程中,雄性配子(精子)和雌性配子(卵子)这两种单倍体细胞融合在一起形成受精卵,受精卵就是二倍体。
受精卵中的一个染色体组来自母亲,另一个染色体组来自父亲。
我需要对另一个观点进行纠正。虽然这对我们要探讨的问题无关紧要,但这个观点非常有意思,因为它能证明每一个单独的染色体组中都包含完整的“模式”代码文本。
在一些例子当中,减数分裂之后并没有立即受精,单倍体细胞(配子)历经许多次细胞有丝分裂,最终产生完整的单倍体个体。雄性蜜蜂就是孤雌生殖产生的个体,从蜂后产下的未受精卵——也就是单倍体的卵发育而来。所以,雄峰是没有父亲的!它所有的体细胞都是单倍体。如果你愿意,你甚至可以叫它“夸张的精子”,实际上正如所有人知道的那样,雄蜂生命中唯一的任务就是产生精子。但这恐怕是一种荒唐的观点。这个例子并不是独一无二的。有很多种植物通过减数分裂产生叫做孢子的单倍体配子,会掉进土壤当中,像种子一样发育成一棵和二倍体植物大小相当的单倍体植物。图5是一种森林中常见的苔藓植物的草图。底部叶状的部分是单倍体植物,称为配子体,它的顶端会长出生殖器官和配子,配子通过相互受精产生普通的二倍体植物,也就是顶端长了孢蒴的光秃的茎。它叫做孢子体,因为它顶端的孢蒴能通过减数分裂产生孢子。孢蒴打开的时候,孢子就会落在土里,长成叶状体,如此往复。这一事件的过程叫做世代交替。如果你愿意,也可以用相同的视角看待普通生物,比如人和动物。不过一般来说,这些生物的“配子体”是短暂的单细胞世代,也就是精子或卵子。我们的身体就是孢子体。我们的“孢子”就是那些“保存”下来的细胞,它们通过减数分裂产生了单细胞的配子世代。(https://www.xing528.com)
个体繁殖过程中重要的、真正决定命运的事件并不是受精,而是减数分裂。我们的一组染色体来源于父亲,另一组染色体来源于母亲。运气也好,命数也罢,都干扰不了这个过程。每个人的遗传都是刚好一半来自父亲,一半来自母亲。而由于一些别的原因,来自一方的遗传往往会比另一方的更明显,这一话题我们会在后面进行讨论(性别本身就是最显而易见的显性例子)。
但是如果你把遗传的来源一直追溯到祖代,情况就不同了。现在我们只讨论来自我父亲的染色体组,特别是其中的一条染色体,比如说5号染色体。这个染色体跟我父亲从他父亲或母亲那里获得的5号染色体中的一个是一模一样的。而我父亲传给我的5号染色体究竟是跟祖父传给他的那条一样,还是跟祖母传给他的那条一样,是由1886年11月我父亲的某个细胞发生的完全随机的减数分裂决定的,概率是一半一半,减数分裂产生的精子几天之后就会携带着这条染色体产生我。我身体中来自父亲的染色体组中的1号、2号、3号……24号染色体都会经历同样的过程,同样的情况也适用于来自母亲的染色体组中的每一条染色体。而且,这48条染色体的分配都是完全独立的。即便已知我父亲遗传给我的5号染色体来自我的祖父约瑟夫·薛定谔,我父亲遗传给我的7号染色体还是有一半可能来源于我祖父,还有一半可能来源于我祖母玛丽·博格纳。
但是因为一些完全随机的因素,一个人的身体当中,祖辈遗传物质的混合比上面讨论的情况更加复杂,因为上面的情况有个假定的前提。说白了就是对于一个特定的染色体来说,它都完整地遗传自祖父或祖母,换句话说,一个染色体在代代相传的过程中是完整不可分割的。然而事实并非如此,或者并不总是这样。在减数分裂中染色体分别进入两个子细胞之前,任何两个“同源”染色体都会彼此接触,并且在接触的时候可能会随机交换一部分,就像图6所示的那样。我们把这个过程叫做“互换”,通过这种机制,分别位于同一个染色体的两部分上的性状就会在孙代发生分离,孙代中一个性状会随祖父,而另一个性状则会随祖母。
左:接触中的两个同源染色体。右:交换和分离以后。
互换的行为并不罕见,也没有非常频繁,但给我们提供了关于染色体上性状相对位置的宝贵信息。要完整地解释这个问题,我们需要用到下一章会提出的一些概念(比如杂合子、显性等);但是这样我们就偏离了这本小册子的主要内容,所以我会简单地指出一些要点。
如果不存在互换现象,那么同一条染色体决定的两个性状总是会一起遗传给下一代,不会出现后代得到了其中一个性状,却没有得到另一个性状的情况。而不在同一条染色体上的两个性状有50%的概率会分离,如果它们位于同一个亲代的同源染色体上,在后代就会分离,因为同源染色体不可能进入同一个子细胞。
这种规律和概率会由于互换现象受到影响。所以通过大量精心设计的繁育实验,仔细统计后代中性状的比例,就能够推断出互换现象发生的概率。这背后的统计学分析基于一个假设的前提,位于同一个染色体上的两个性状距离越近,它们的“连锁”就越不容易受到互换的影响。由于距离的两个性状之间发生交换的概率小,而靠近染色体两端的性状容易发生互换从而产生分离。(同一个亲代的同源染色体上的性状自由组合也遵循同样的规律)按照这一原理,通过对“连锁统计数据”进行分析,我们就能够绘制出每个染色体上的“性状图”。
这种设想已经完全被证实了。人们已经在一些动物上进行了充分的实验验证(主要但不限于果蝇),进行验证的性状实际上分成了许多不同的、相互独立的组,这些不同的组代表不同的染色体(果蝇有4对染色体)。对于每一组性状,通过对任何两个性状之间的连锁程度进行量化分析,就能够绘制出性状排列位置的线形图,这些性状实际上也几乎是按照线性排列的,这一结论与染色体的棒状结构也是相符的。
当然,上面描绘出的遗传机制的草图仍然不够复杂,甚至可以说有点简单。因为我们还没有解释对性状的理解。将一种生物的模式分割成离散的“性状”,似乎理由并不充分,而且也是不可能的,因为生物的模式是统一的,是一个“整体”。
现在,对于任何生物来说,我们所说的性状是指,一对亲本个体在某一个具体方面有差异(比如,一个是蓝眼睛,一个是棕眼睛),而后代这方面的特征与父母之一是一样的。我们在染色体上定位的实际上是产生这种差异的位置。(生物学术语称之为“基因座”,或者说,从其背后的物质结构,也就是“基因”。)性状的差异在我看来才是真正的基本概念,而性状本身并不是,虽然这种说法本身用词不够准确,逻辑也说不通。
性状的差异本身是离散的,下一章我们讨论到突变时就会发现这一点,而且我希望通过下一章的内容,我们目前对遗传机制干巴巴的描绘能够丰富多彩起来。
我们刚刚引入的基因这个概念,将其定义为:一种特定的遗传特征对应的理论物质载体。我们现在必须强调,与我们的探讨密切相关的两点。第一点是物质载体的体积,更准确地说是它的最大体积,换句话说,我们确定基因位置时能够精确到多小的体积范围。第二点是基因的永久性,这一点是从遗传规律的持久性推断而来的。对于体积问题,人们通过两种不同的方法进行了估计,其中一种基于遗传学的证据(繁育实验),另一种则基于细胞学证据(在显微镜下直接观察)。
从原则上讲,第一种方法非常简单。通过上面描述的方法,定位一条特定染色体上大量不同(宏观尺度)性状(比如以果蝇为对象)后,染色体长度除以性状的数量,再乘以横截面面积,就得到了我们所需要的基因体积的估计数。当然,我们只认为能够偶尔发生互换的性状是不同的,这样它们的(显微或分子)结构就不可能是一样的。另外,这种估计方法只能得出基因体积的上限,因为随着研究的深入,通过这种遗传分析确认的性状数量也在不断增加。
而另一种估计方法虽然建立在显微观察的基础上,但事实上不如上一种方法直接。由于某些原因,果蝇身体中的某些细胞(即唾液腺细胞)体积非常大,它们的染色体也非常大。你能够看到染色体纤维上面分布着紧密排列的深色横向条纹。C.D.达林顿指出,这些条纹虽然在数量上比繁育实验确定的遗传性状大得多(他使用的数字是2000),但是二者仍然处于同一个数量级上。他倾向于将这些条纹视为真正的基因(或者基因之间的间隔)。用正常大小细胞中染色体的长度除以这个数字(2000),他发现基因的体积大约与边长为300埃的立方体相当。考虑到这两种估计方法的粗糙,我们基本上可以认为这两种估计方法得到的数字是差不多的。
后面我们会详细讨论统计物理学与我能想到的所有事实之间的联系,或者我应该说,这些事实与统计物理学应用于活体细胞之间的关系。但请注意一个事实,液体或固体状态下,300埃只有150个左右的原子距离,所以一个基因包含的原子数量不过一百万或几百万个。这一数字太小了(从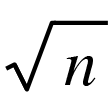 法则角度来看),从统计物理学的角度来看,也就是说从物理学的角度来看,根本不可能产生有秩序、有规律的行为。即便所有这些原子起的作用都是一样的,就像它们在气体或液滴当中那样,这个数字也太小了。而且基因肯定不会像质地均匀的液滴。它可能是一个巨大的蛋白质分子,其中的每个原子、每个原子团、每个杂环都有各自的作用,或多或少与其他相似的原子、原子团或杂环有些不同。无论如何,这是前沿遗传学家霍尔丹和达林顿的观点,我们很快就会讲到几乎可以证明这一观点的遗传学实验。
法则角度来看),从统计物理学的角度来看,也就是说从物理学的角度来看,根本不可能产生有秩序、有规律的行为。即便所有这些原子起的作用都是一样的,就像它们在气体或液滴当中那样,这个数字也太小了。而且基因肯定不会像质地均匀的液滴。它可能是一个巨大的蛋白质分子,其中的每个原子、每个原子团、每个杂环都有各自的作用,或多或少与其他相似的原子、原子团或杂环有些不同。无论如何,这是前沿遗传学家霍尔丹和达林顿的观点,我们很快就会讲到几乎可以证明这一观点的遗传学实验。
现在我们讨论第二个与我们的主题密切相关的问题。遗传性状的稳定性究竟有多强,这种稳定性与承载遗传性状的物质结构又有多大关系呢?
其实不需要进行任何特殊的研究,我们就可以回答这个问题。
我们能够讨论遗传性状本身就证明,我们几乎完全认可了它的稳定性。我们必须记住,父母遗传给孩子的并不仅仅是某个特点,比如鹰钩鼻、短手指、易患风湿、血友病、二色性色盲等等。我们也许能够方便地选取这些特点用于研究遗传定律。但是这些特点实际上是产生“表现型”的完整(四维)模式,是个体可见的、明显的本性,它们由结合成受精卵的两个细胞的物质结构携带,经过数千年的代际传播也不会发生显著变化,不过在几万年的时间当中这种稳定性就难说了。
这可以说是个奇迹,不过还有一个更伟大的奇迹。这两个奇迹虽然密切相关,却在不同的层面上。我说的另一个奇迹是指:虽然我们人类的存在全部建立在这种奇迹般的相互作用上,但是能够掌握获取这个奇迹相关知识的能力。我认为这些知识也许能够让我们对第一个奇迹产生近乎完整的了解。
但第二个奇迹则可能超出人类的理解范围。
[1]实际上人类有46条染色体,此为薛定谔演讲中的错误,后面也应该写作2×23。下文同理。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。