



物质在生物体内经过一系列连续的氧化还原反应,逐步分解并释放能量的过程,这个过程也称为生物氧化,是一个产能代谢过程。在生物氧化过程中释放的能量可被微生物直接利用,也可通过能量转换储存在高能化合物(如ATP)中,以便逐步被利用,还有部分能量以热的形式被释放到环境中。不同类型微生物进行生物氧化所利用的物质是不同的,异养微生物利用有机物,自养微生物则利用无机物,通过生物氧化来进行产能代谢。
一、化能异养型微生物的能量代谢
(一)化能异养型微生物ATP的产生
ATP是生物细胞内的通用能源,贮存能量时,ADP(二磷酸腺苷)结合1分子磷酸生成ATP;供能时,ATP末端高能键水解脱去1分子磷酸,生成ADP释放能量。
ADP+Pi+35kJ/mol→ATP
除ATP外,还有一些重要的高能化合物如乙酰磷酸、PEP(磷酸烯醇式丙酮酸)等,这些物质水解释放的能量贮存在ATP中。ATP贮存的能量又能在激酶作用下转移至其他化合物。ATP的形成按能量来源分为光合磷酸化和氧化磷酸化两种方式。光合磷酸化是光能营养微生物形成ATP的途径;氧化磷酸化是化能微生物形成ATP的途径,根据电子传递链的有无,可分为底物水平磷酸化和电子传递磷酸化两种类型。
1.底物水平磷酸化
物质在生物氧化过程中,生成一些含有高能键的化合物,而这些化合物可直接偶联ATP或GTP(三磷酸鸟苷)的合成,这种产生ATP等高能根子的方式称为底物水平磷酸化。例如发生底物水平磷酸化的几种重要反应。
底物在生物氧化中脱下的电子或氢通过酶促反应直接交给底物自身的氧化产物,同时释放出的能量交给ADP,形成ATP。底物水平的磷酸化是微生物在发酵中产生ATP的唯一方式,在呼吸过程中也有存在。
2.电子传递磷酸化
底物在生物氧化过程中释放的电子通过电子传递链传递到氧或其他氧化型物质,同时形成ATP的过程称为电子传递磷酸化。电子传递磷酸化的核心为电子传递链(electron transport chain,ETC),其组成成员主要有泛醌(辅酶Q)、NAD(烟酰胺腺嘌呤二核苷酸)与NADP(烟酰胺腺嘌呤二核苷酸磷酸)、FAD(黄素腺嘌呤二核苷酸)和FMN(黄素腺嘌呤单核苷酸)、铁硫蛋白及细胞色素5类物质。E TC是由若干个氢和电子传递体按氧化还原电位高低顺序排列而构成的一条链,流动的电子通过呼吸链时做功,逐步释放能量形成ATP。
(二)化能异养型微生物的生物氧化
1.发酵
发酵是指微生物细胞将有机物氧化释放的电子直接交给底物本身未完全氧化的某种中间产物,同时释放能量并产生各种不同的代谢产物。在发酵条件下有机化合物只是部分地被氧化,因此,只释放出一小部分的能量。发酵过程的氧化是与有机物的还原偶联在一起的。被还原的有机物来自于初始发酵的分解代谢,即不需要外界提供电子受体。
发酵的种类有很多,可发酵的底物有碳水化合物、有机酸、氨基酸等,其中以微生物发酵葡萄糖最为重要。生物体内葡萄糖被降解成丙酮酸的过程称为糖酵解,主要分为四种途径:EMP途径、HMP途径、ED途径、磷酸解酮酶途径。
(1)EMP途径(Embden—Meyerhof pathway) 以葡萄糖为起始底物,丙酮酸为终产物。整个EMP途径大致可分为两个阶段。第一阶段可认为是不涉及氧化还原反应及能量释放的准备阶段,只是生成两分子的主要中间代谢产物:3—磷酸—甘油醛。第二阶段发生氧化还原反应,合成ATP并形成两分子的丙酮酸,如图6-1所示。
图6-1 EMP途径
a.预备性反应,消耗ATP,生成甘油醛—3—磷酸
b.氧化还原反应,形成ATP和产生发酵产物
EMP途径可为微生物的生理活动提供ATP和NADH,其中间产物又可为微生物的合成代谢提供碳骨架,并在一定条件下可逆转合成多糖。
(2)HMP途径 HMP途径是从6—磷酸葡萄糖酸开始的,即在单磷酸己糖基础上开始降解的,故称为单磷酸己糖途径。HMP途径与EMP途径有着密切的关系,因为HMP途径中的3—磷酸甘油醛可以进入EMP,因此该途径又可称为磷酸戊糖支路。HMP途径的一个循环的最终结果是一分子6—磷酸葡萄糖转变成一分子3—磷酸甘油醛,三分子CO2和六分子NADPH。
一般认为HMP途径不是产能途径,而是为生物合成提供大量的还原力(NAD-PH)和中间代谢产物。
大多数好氧和兼性厌氧微生物中都有HMP途径,而且在同一微生物中往往同时存在EMP和HMP途径,单独具有EMP或HMP途径的微生物较少见。
(3)ED途径在ED途径中,6—磷酸葡萄糖首先脱氢产生6—磷酸葡萄糖酸,接着在脱水酶和醛缩酶的作用下,产生一分子3—磷酸甘油醛和一分子丙酮酸。然后3—磷酸甘油醛进入EMP途径转变成丙酮酸。一分子葡萄糖经ED途径最后生成两分子丙酮酸,一分子ATP,一分子NADPH和NADH。
ED途径在细菌中,尤其是在革兰氏阴性菌中分布较广,特别是假单胞菌和固氮菌的某些菌株较多存在。ED途径可不依赖于EMP和HMP途径而单独存在,但对于靠底物水平磷酸化获得ATP的厌氧菌而言,ED途径不如EMP途径经济。
(4)磷酸解酮酶途径 磷酸解酮酶途径是明串珠菌在进行异型乳酸发酵过程中分解己糖和戊糖的途径。该途径的特征性酶是磷酸解酮酶,根据解酮酶的不同。把具有磷酸戊糖解酮酶的称为PK途径,把具有磷酸己糖解酮酶的叫HK途径。(https://www.xing528.com)
在糖酵解过程中生成的丙酮酸可被进一步代谢。丙酮酸由脱羧酶催化形成乙醛和二氧化碳,乙醛在乙醇脱氢酶的作用下,被NADH还原为乙醇。这种氧化作用不彻底,只释放出部分能量,而大部分能量还贮存在乙醇中。发酵作用的总反应式如下:
C6H12O6+2ADP+2Pi→2CH3CH2OH+2CO2+2ATP
各种微生物都能进行发酵作用。在无氧条件下,不同的微生物分解丙酮酸后会积累不同的代谢产物。目前发现多种微生物可以发酵葡萄糖产生乙醇,能进行乙醇发酵的微生物包括酵母菌、根霉、曲霉和某些细菌。许多厌氧菌主要靠发酵作用获取能量。好养微生物在进行有氧呼吸过程中,也要先经过糖酵解阶段产生丙酮酸,然后进入三羧酸循环,将底物彻底氧化成二氧化碳和水。
2.呼吸作用
微生物在降解底物的过程中,将释放出的电子交给NAD(P)+、FAD或FMN等电子载体,再经电子传递系统传给外源电子受体,从而生成水或其他还原型产物并释放出能量的过程,称为呼吸作用。其中,以分子氧作为最终电子受体的称为有氧呼吸,以氧化型化合物作为最终电子受体的称为无氧呼吸。
呼吸作用与发酵作用的根本区别在于:电子载体不是将电子直接传递给底物降解的中间产物,而是交给电子传递系统,逐步释放出能量后再交给最终电子受体。
(1)有氧呼吸 葡萄糖经过糖酵解作用形成丙酮酸,在发酵过程中,丙酮酸在厌氧条件下转变成不同的发酵产物;而在有氧呼吸过程中,丙酮酸进入三羧酸循环(tricarboxylic acid cycle,简称TCA循环),被彻底氧化生成CO2和水,同时释放大量能量,如图6-2所示。
(2)无氧呼吸 某些厌氧和兼性厌氧微生物在无氧条件下进行无氧呼吸。无氧呼吸的最终电子受体不是氧,而是像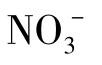 、
、 、
、 、
、 、CO2等这类外源受体。无氧呼吸也需要细胞色素等电子传递体,并在能量分级释放过程中伴随有磷酸化作用,也能产生较多的能量用于生命活动。但由于部分能量随电子转移传给最终电子受体,所以生成的能量不如有氧呼吸产生的多。
、CO2等这类外源受体。无氧呼吸也需要细胞色素等电子传递体,并在能量分级释放过程中伴随有磷酸化作用,也能产生较多的能量用于生命活动。但由于部分能量随电子转移传给最终电子受体,所以生成的能量不如有氧呼吸产生的多。
图6-2 三羧酸循环
二、自养微生物的生物氧化与产能
(一)化能自养菌的生物氧化与产能
一些微生物可以从氧化无机物获得能量,同化合成细胞物质,这类细菌称为化能自养微生物。它们在无机能源氧化过程中通过氧化磷酸化产生ATP。化能自养菌是一类以还原性无机物如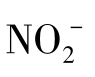 盐、硫化氢、Fe2+或氢气等作为电子供体并利用这些无机物的氧化释放出能量,以CO2或碳酸盐为唯一或主要碳源合成细胞物质的微生物。此类微生物主要有硝化菌群、硫细菌、铁细菌和氢细菌四大类。化能自养微生物的生物氧化包含以下几种类型。
盐、硫化氢、Fe2+或氢气等作为电子供体并利用这些无机物的氧化释放出能量,以CO2或碳酸盐为唯一或主要碳源合成细胞物质的微生物。此类微生物主要有硝化菌群、硫细菌、铁细菌和氢细菌四大类。化能自养微生物的生物氧化包含以下几种类型。
1.氨的氧化
NH3同亚硝酸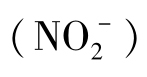 是可以用作能源的最普通的无机氮化合物,能被硝化细菌所氧化。硝化细菌可分为两个亚群:亚硝化细菌和硝化细菌。氨氧化的过程分为两个阶段,先由亚硝化细菌将氨氧化为亚硝酸,再由硝化细菌将亚硝酸氧化为硝酸。由氨氧化为硝酸是通过这两类细菌依次进行的。
是可以用作能源的最普通的无机氮化合物,能被硝化细菌所氧化。硝化细菌可分为两个亚群:亚硝化细菌和硝化细菌。氨氧化的过程分为两个阶段,先由亚硝化细菌将氨氧化为亚硝酸,再由硝化细菌将亚硝酸氧化为硝酸。由氨氧化为硝酸是通过这两类细菌依次进行的。
硝化细菌都是一些专性好氧的革兰氏阳性细菌,以分子氧为最终电子受体,且大多数是专性无机营养型。它们的细胞都具有复杂的膜内褶结构,这有利于增加细胞的代谢能力。硝化细菌无芽孢,多数为二分裂殖,生长缓慢,平均代时在10h以上,分布非常广泛。
2.硫的氧化
硫细菌是一群能够利用一种或多种还原态或部分还原态的硫化合物(包括硫化物、元素硫、硫代硫酸盐、多硫酸盐和亚硫酸盐)氧化释放能量生长的细菌。多数硫化细菌为专性化能自养,专性好氧,少数为兼性化能自养与兼性厌氧。H2S首先被氧化成元素硫,随之被硫氧化酶和细胞色素系统氧化成亚硫酸盐,放出的电子在传递过程中可以偶联产生四个ATP。亚硫酸盐的氧化可分为两条途径,一是直接氧化成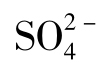 的途径,由亚硫酸盐—细胞色素C还原酶和末端细胞色素系统催化,产生一个ATP;二是经磷酸腺苷硫酸的氧化途径,每氧化一分子
的途径,由亚硫酸盐—细胞色素C还原酶和末端细胞色素系统催化,产生一个ATP;二是经磷酸腺苷硫酸的氧化途径,每氧化一分子 产生2.5个ATP。
产生2.5个ATP。
3.铁的氧化
从亚铁到高铁状态的铁的氧化,对于少数细菌来说也是一种产能反应,但从这种氧化中只有少量的能量可以被利用。亚铁的氧化仅在嗜酸性的氧化亚铁硫杆菌中进行了较为详细的研究。在低pH环境中这种菌能利用亚铁放出的能量生长。在该菌的呼吸链中发现了一种含铜蛋白质,它与几种细胞色素c和一种细胞色素a1氧化酶构成电子传递链。虽然电子传递过程中的放能部位和放出有效能的多少还有待研究,但已知在电子传递到氧的过程中细胞质内有质子消耗,从而驱动ATP的合成。
4.氢的氧化
氢细菌都是一些呈革兰氏阴性的兼性化能自氧菌。它们能利用分子氢氧化产生的能量同化CO2,也能利用其它有机物生长。氢细菌的细胞膜上有泛醌、维生素K2及细胞色素等呼吸链组分。在该菌中,电子直接从氢传递给电子传递系统,电子在呼吸链传递过程中产生ATP。在多数氢细菌中有两种与氢的氧化有关的酶。一种是位于壁膜间隙或结合在细胞质膜上的不需NAD+的颗粒状氧化酶,它能够催化以下反应:
H2→2H++2e-
该酶在氧化氢并通过电子传递系统传递电子的过程中,可驱动质子的跨膜运输,形成跨膜质子梯度为ATP的合成提供动力;另一种是可溶性氢化酶,它能催化氢的氧化,而使NAD+还原的反应。所生成的NADH主要用于CO2的还原。
(二)光能自养菌的生物氧化与产能
光合作用是自然界一个极其重要的生物学过程,其实质是通过光合磷酸化将光能转变成化学能,以用于从CO2合成细胞物质。光能自养菌是一类能以CO2为唯一碳源或主要碳源并利用光能进行生长的微生物,如藻类、蓝细菌和嗜盐细菌等。光合细菌主要通过环式光合磷酸化作用将光能转变为化学能,产生ATP。在光合细菌中,吸收光量子而被激活的细菌叶绿素释放出高能电子,叶绿素分析即带正电荷。所释放出来的高能电子顺序通过铁氧还蛋白、辅酶Q、细胞色素b和c,再返回到带正电荷的细菌叶绿素分子。在辅酶Q将电子传递给细胞色素c的过程中,造成了质子的跨膜运动,为ATP的合成提供了能量。在这个电子循环传递过程中,光能转变为化学能,故称为环式光合磷酸化。环式光合磷酸化可在厌氧条件下进行,产物只有ATP,无NADP(H),也不产生分子氧。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。