



四、林木分子生物学
林木分子生物学研究,实际上是指林木基因工程研究,它借助于现代生物技术,结合林木特点,建立起自己的测试方法并在以下几方面得到开展。
1.林木遗传图谱
目前,林木遗传图谱构建是林木分子遗传学领域研究的重要新课题。它利用分子标记技术包括RFLP、RADP、AFIP、SSR等交叉使用,揭示林木物种的遗传性状并制出图谱。这是一项繁琐而复杂的工作,因为技术遗传背景材料知之甚少,给遗传图谱构建带来一定的难度和不便。但大多林木都可以进行无性繁殖而重要的经济性状皆为数量性状,这样就比较易于进行QTLS(数量性状基因位点)定位的研究。
在针叶树中,由于大配子体为单倍体,其显性与非显性标记具有相同的遗传行为,因此,可以利用单株树的种子进行遗传作图和QTLS定位,所及控制数量性状的基因、数目、在染色体上的位置以及各位点贡献的大小和基因间相互关系。现已定位的QTLS包括生长、木材密度、抗干旱胁迫、抗病等,涉及树种有火炬松、湿地松、辐射松、杨树、杉木、马尾松、桉树等造林树种,还有苹果、柑橘、桃树等果树。
在以上工作中,由南京林业大学王明庥主持的“杉木和杨树遗传图谱构建和数量性状基因定位”课题,经过多年的努力,取得了突破性进展(尹伟伦,胡建军,2005);构建了5张遗传连锁图谱,包括响叶杨、银白杨的分子标记连锁图谱,美洲黑杨、欧美杨AFLP指纹图谱,尾叶桉和细叶桉RAPD标记图谱以及杉木和马尾松遗传图谱;并对树种作图群体的性别分化、木材密度和生根等性状进行了QTL定位分析,探索了林木复杂性状基因定位的新策略。应该说,目前建筑的若干种的遗传连锁图谱还只是一个框架图,大多没有和染色体相对应。然而,作为木本模式植物杨树(Populus trichoarpa)全基因组框架图的完成,并配以杨树功能基因组学,基因表达研究必将为林木基因工程研究创造了条件。
分子遗传标记是建于核酸水平上具有相差异的等位基因,通过分子标记技术的连锁图谱的构建,来揭示林木优良性状的遗传背景,加速林木性遗传改良进程而不要对遗传材料进行特殊处理的一种方法。目前,常用的DNA分子标记主要有以下5种。
(1)RFLP(限制性片段长度多态性)是最早发展的分子标记。其基本原理是利用限制性内切酶酶切不同生物个体基因组DNA后,转移于支持膜上与放射性核素(32P)的探针进行Southern杂交,从而显示与探针含同源顺序的酶切片段在长度上的差异,检测不同遗传位点的等位变异(多态性)。RFLP是一种共显性标记,非等位RFLP标记之间不存在上位效应,结果稳定可靠,重复性好,因而用于构建表达图谱、分析群体内和群体间遗传变异度及亲缘关系等。
(2)RAPD(随机扩增片段长度多态性)标记技术,通常以10个碱基的寡核苷酸序列为引物,利用对基因组DNA随机扩增来得到多态性图谱。扩增产物通过琼脂糖凝胶电泳等分离,经EB染色后,在紫外透射仪上检测扩增DNA片段的多态性,并对此基因组DNA进行多态性分析,进行遗传作图品种鉴定。
(3)SSR(简单重复序列)标记,又称微卫星DNA标记,是指以几个核苷酸(1~6个)为单位,多次串联重复的DNA序列。广泛分布于真核生物基因组中,大约每隔10~50bp就有一个微卫星,由于重复次数和重复程序的不完全而造成每一个位点的多态性。SSR两端有一段保守的DNA序列,通过它可以设计一对互补的寡核酸引物,经电泳检测其多态性。
(4)AFLP(扩增片段长度多态性)标记,结合了RFLP和RAPD的优点,通过对基因组DNA酶切片段的选择性扩增来检测DNA酶切片段长度多态性。由于AFLP扩增可使某一品种出现特定的DNA谱带,而在另一品种中可能无此谱带产生,因此这种通过引物诱导及DNA扩增后得到的DNA多态性可作为一种分子标记。
(5)ISSR(inter-simple sequence repeat)是Zietkeiw itcz等于1994年发展起来的一种微卫星基础上的分子标记。其基本原理是:用锚定的微卫星DNA为引物,即在SSR序列的3′端或5′端加上2~4个随机核苷酸,在PCR反应中,锚定引物可引起特定位点退火,导致与锚定引物互补的间隔不太大的重复序列间DNA片段进行PCR扩增。所扩增的inter SSR区域的多个条带通过聚丙烯酞胺凝胶电泳得以分辨,扩增谱带多为显性表现。ISSR引物的开发不像SSR引物那样需测序获得SSR两侧的单拷贝序列,开发费用降低。与SSR标记相比,ISSR引物可以在不同的物种间通用,不像SSR标记一样具有较强的种特异性;与RAPD和RFLP相比,ISSR揭示的多态性较高,可获得几倍于RAPD的信息量,精确度几乎可与RFLP相媲美,检测非常方便,因而是一种非常有发展前途的分子标记。目前,ISSR标记已广泛应用于植物品种鉴定、遗传作图、基因定位、遗传多样性、进化及分子生态学研究中。
还需指出,近年来植物功能基因组学研究技术包括表达序列标签(EST)、基因表达的系列分析(SAGE)、DNA微阵列、反向遗传学发展已在林木中应用通过基因克隆建筑数据库、鉴定与分析基因序列、片段在特定组织中表达。有助于阐述林木基因组的结构与功能,并为林木分子改良育种提供有效依据。
2.果木开花的基因控制
高等植物在萌发之后需要经历一定时间的营养生长,即童期(juvanility),才能进入生殖发育阶段。控制植物生殖转变调节童期的基因主要有花序分生组织特异基因、花分生组织特异基因、花器官分生组织特异基因,对其几种模式植物,如拟南芥、金鱼草、烟草进行了充分的研究与揭示。
然而,多年生果木植物需要经历一个较长的童期(4~20年),才能开花结果,这一特性极大地限制了有经济价值的果木品种的遗传改良。近几年,根据开花调节基因的相对同一性和保守性,从木本植物上分离克隆了许多基因,包括花序分生组织特异基因(TFL)同源基因、花分生组织特异基因(LFY、APL)同源基因、花器官特异(MADS-box)基因及同源基因(APL、DEF、AG),以研究异位表达开花调节基因诱导开花和缩短童期的作用。
TFL是拟南芥中控制分生组织中花基因表达的一个基因,它的表达受到LFY、AG的负调控。Kotoda等(2003)从苹果上克隆了LFY的同源基因MdTFL,对其表达模式进行研究,表明该基因在花诱导前两周的萼片与成熟叶片表达较强。将反义MdTFL基因转入苹果,结果表明花期提早2~3年。LFY基因是控制植物从营养状态转向生随状态过程中起重要调节近代作用的转录因子,是开花启动所必需的。LFY、APL的同源基因也从桉树、辐射松、银杏、白杨中被分离克隆并在花组织和营养组织中进行了表达,对开花也起到一定的调节作用。
按照花器官ABC模式,将MADS-box基因分为三种类型:A类决定萼片形成,如APETALA1基因;B类和A类一起控制花瓣形成,如AP3/P1(拟南芥)、DEF/G10(金鱼草);C类和B类共同作用决定雄蕊分化,C类单独表达心皮的形成,如AG。近几年,木本植物MADS-box基因的分离克隆较多,主要集中在杨树、桦树、桉树、苹果、柑橘、猕猴桃上,工作在逐步开展。尽管植物开花调节已被大量分离和克隆而且转基因可以大大缩短木本的童期,但是,果木植物的开花生理机制是很复杂的,远远未被揭示;加之,多数木本植物的组织培养,离体再生和基因转化技术还存在一定难度,所以,开花调节基因的转化研究在木本上的应用还不多。
然而,白桦(betula)幼树强化诱导促进开花结实是一个特殊的例子,值得一提,早在1959年,美国Longman和Wareing发现桦树幼树苗的连续生长可以刺激开花的生理现象。直到1967年,芬兰树木育种基金会工作者建立塑料大棚对1年生疣枝桦(B.Varrucosa)实生苗进行连续生长的控制,于1969年获得一批种子。随之对欧洲白桦(B.Pendula)进行提早开花结实选育种研究,并于1972年第一座生产欧洲白桦塑料大棚种子园建立,以满足生产需求(Hannu 1991)。自此以后这项研究工作引起了林木育种学和植物生理学工作者的关注。
根据杨传平等(2004)报道,东北林业大学林学院在20世纪90年代,从芬兰获得有关设备,建立了室内强化育种园。在9年42次的前期试验的基础上,以1年生超级白桦苗为材料,通过5项配套强化措施,包括适量CO2浓度、适当光照强度、适时绞缢、适时催化素喷施和适中温湿度的控制的处理,实现了野外条件17~20年开花结实的白桦,2~3年开花结实,4~5年规模结实。这如何解释白桦幼苗强化培育提早开花结实的遗传生理机理呢?通过单项优化处理在生理学上多少是有作用的,但不能改变遗传的开花习性。再从模式植物拟南芥(arabidopsis)成花分子机理研究所取得的进展上看,不是克隆某类成花茎因转入其他植物就能开花的。所以说,高等植物成花是一个非常复杂的过程。分子生物学成花基因和环境强化成化条件都需要深入研究,还应交叉开展有助于理论与生产结合,实现从特殊性到普遍性。(https://www.xing528.com)
3.植物的抗冻基因
1985年Guy等发现植物在低温驯化过程中,基因表达会发生改变,由此推动了抗冻基因的研究。随之,植物抗冻机制的研究发现了抗冻蛋白的功能。抗冻蛋白已在抗冻植物,如冬小麦、胡萝卜、黑麦草及鱼、昆虫和细菌中被发现。
1992年,加拿大M.Griffith等首先在冬黑麦中发现了植物抗冻蛋白(AFPs),1993年Duman等从越冬的欧白英(Solnum dulcamara)枝条中分离出一分子量为67kD的抗冻蛋白(AFGP)。我国魏令波等从强抗冻植物沙冬青(Ammopiptanthusmongolicus)叶片分离出热稳定的抗冻糖蛋白,分子量为40kD,当浓度为20g/L时,其热带活性为0.9℃。1999年Michael等从桃树的树皮中提取到一种脱水蛋白PCA60,这是第一次发现具有脱水蛋白特性的AFPs。卢存福等(2000)从耐寒的高山植物唐古特红景天低温悬浮细胞液中检测AFPs,这也是AFPs主要在植物质外体系统分布的间接证据。他们还选择了耐干旱、盐碱、寒冷、抗风沙,能在夏季酷热,冬季“寒极”的“三北”恶劣环境下生长的胡杨为实验材料,建立了稳定的胡杨悬浮细胞培养体系,并首次从木本植物悬浮培养细胞获得部分纯化的抗冻蛋白,用差示扫描的量热法,测定低温诱导25天细胞分泌蛋白的热滞活性(THA)值,发现和鱼1-型抗冻蛋白(Typ-1 AFP)有相似的差热分析曲线,测得蛋白浓度为1.0mg/ml时,热滞活性值约0.67℃;对66kD、36kD、22kD多肽分离纯化、抗冻活性测定及抗冻培育工作尚在进行之中。
近几年来,北京林业大学林善技和张志毅(2004)选取具有极强抗旱抗冻性的甜杨(P.suaveloens)为材料,开展抗冻特异蛋白的氨基酸组分及含量分抑而获得了相关的氨基酸序列。在此基础上以所获的氨基酸序列为探针,从甜杨总RNA中克隆出(图10-1)。试验将甜杨60kD抗冻蛋白氨基酸组分含量与其他抗冻蛋白进行比较,发现它与沙冬青40kD抗冻性以及Sd-67相似,但在Gly和Ser含量上仍有明显差异;甜杨60kD的热滞性为2.2℃,较鱼类(0.7- 1.5℃)、桃树、冬黑麦和沙冬青等抗冻性蛋白(0.5- 0.9)的热滞活性值要高一些。这意味着甜杨抗冻性蛋白与其他灌木和草本植物有区别,它可能属于一种高热滞活性的抗冻蛋白。
从代谢角度看,葡萄糖-6-磷酸脱氢酸(G6PDH)是戊糖磷酸循环(PPP)的关键性调控酶。Sagisava(1985)研究发现,在低温贮藏下杨树枝条内G6PDH活性明显增加,而这种酶的钝化或失活会导致枝条冻害的发生。目前已在油菜、大麦、玉米、豌豆等植物低温锻炼中发现了G6PDH活性和NADPH2水平的提高。林善技等(2004)将甜杨幼苗处于- 20℃下低温锻炼获得了同样的结果,并对G6PDH酶进行了纯化分析。利用KT-PCR技术从甜杨总RNA中扩增出长度为1200bp的G6PDH基因并初步确定甜杨PsG6PDH所编码基因可能是一种与抗冻有关的基因,G6PDH可能是甜杨抗御冰冻低温的一种重要生理活性物质。这也把抗冻生理机制与呼吸代谢PPP途径相联系。
4.林木转基因抗虫性培育
植物抗虫基因工程是现代生物技术领域的重要成果之一,林木转基因难度较大,但也取得了重大进展。自从1987年Fillatti等首次利用农杆菌对银白杨X大齿杨进行转化,获得第一株林木转基因植株以来,此方面的研究不断取得突破,如McCown等(1991)获得了抗舞毒蛾和天幕毛虫的银白杨X大齿杨的转基因植株。
我国在20世纪末和21世纪初营造了以亿亩计的杨树速生用材林基地,但易遭食叶虫如杨尺蠖和舞毒蛾、扇羽蛾等为害。为此,自1998年起,中国林科院林业研究所和中科院微生物研究所合作开展抗虫转基因杨树培育,将苏云金杆菌(Bt)杀虫蛋白基因转入欧洲黑杨,已筛选出一批有Bt杀虫蛋白基因整合至染色体的杨树,经抗虫生物测定证实,杨尺蠖和舞毒蛾的死亡率可达80%~100%(田颖川,1993)。
然而,研究表明Bt杀虫结晶蛋白抗虫谱较窄,害虫易对杀虫结晶蛋白产生耐受性,致使转基因植株的抗虫能力不强,抗虫性不持久。于是设法把2个或3个不同类型的抗虫基因同时导入细胞,能在很大程度上延缓昆虫抗性的发生。所以,他们在以往研究基础上,通过农杆菌的Ti质粒的双元载体将这2个抗虫机制不同的基因同时导入杨树(欧洲黑杨),通过这2个杀虫基因(即Bt基因和蛋白本科抑制剂基因),从而获得抗虫能力强且稳定的转基因杨树。
以上实验,经PCR检测和Southern杂交分析证明,最终获得既含有Bt基因又含有蛋酶抑制剂基因的转基因杨树植株。利用这种杨树饲喂舞毒蛾幼虫的杀虫试验结果表明,转基因杨树具有明显的杀虫活性,同时表明含有Bt基因和蛋白酶抑制剂基因双基因的植株其抗虫能力明显高于仅含有Bt基因的植株。这为探索培育高抗虫性杨树开辟了一条新途径(李明亮等,2002)。此外,东北林业大学遗传育种学科以白桦为材料开展了转基因对舞毒蛾的抗性研究也取得了进展(王志英詹亚光等,2007)
5.林木木质素基因工程改良培育
木质素(lignin)是维管植物的重要组成成分,但木质素却是现代纸浆和造纸工业的一大障碍。因此,研究木质素的合成途径,通过基因工程途径降低木质素含量和改变木质素组成,对提高纸浆得率,改善环境有积极作用。木质素不是多糖,它是由苯基丙烷衍生物的单体构成的聚合物。它的前体是对-香豆素、阿魏酸和芥子酸,经葡糖基化分别转化为对-香豆醇、松柏醇和芥子醇脱氢聚合成木质素(H型、G型和S型木质素)。木质素合量可达18%~38%,主要分布在纤维、导管和管胞中。
目前,人们认同木质单体合成途径有三条,即莽草酸途径、苯丙氨酸途径和木质素合成特异途径,有10多种酶参与木质素单体合成调控(图10-1)。这些酶大多被克隆,为基因调控植物木质素的研究奠定了基础。苯丙氨酸途径是由PAL催化反应开始,PAL是连接莽草酸和苯丙氨酸途径的关键酶。许多研究发现,无论是正义或是反义抑制烟草中PAL活,都会导致木质素含量降低,但是伴有植物的非正常生长。
C4H的作用是催化肉桂酸向香豆酸的转化,该酶活性受抑制后,转基因植株木质素含量减少,植物生长无异常现象,S/G比值降低。4CL是连接苯丙氨酸途径与木质素特异合成途径的关键酶,也是催化羟基因桂酸形成G或S型单体途径中的主要酶。Hu等(1999)从白杨中克隆到2个4CL基因Pt4CL1和Pt4CL2,通过对杨树杂种的Pt4CL1的反义抑制,转基因植株的木质素含量降低45%,纤维素则补偿性地增加了15%,叶、根和茎生长速率提高。贾彩红等(2004)利用根癌农杆菌介导法将反义4CL基因转入三倍体毛白杨(P.tomentosa),转基因植株茎秆剥皮后呈现程度不等的红褐色,而对照为白色。转基因株系木质素含量比对照下降41.73%,而总纤维素含量无明显差别。
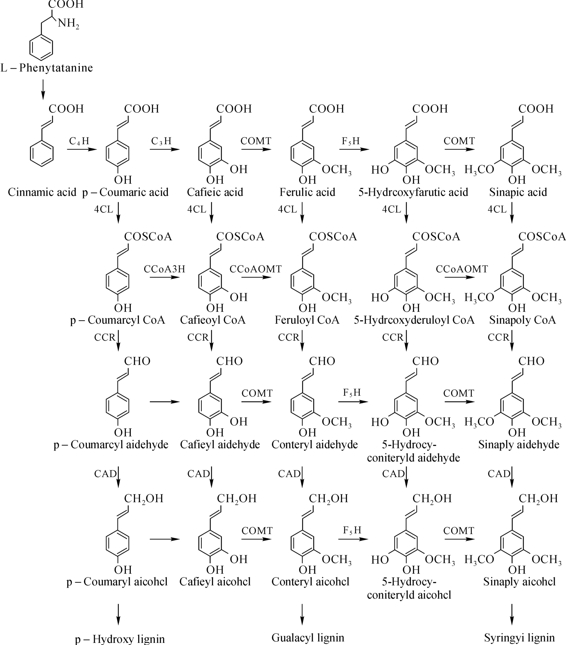
图10-1 木质单体生物合成途径
PAL—苯丙氨酸裂解酶;C4H—肉桂酸-4-、羧基化酶;C3H—香豆素-3-羧化酶;COMT—咖啡酸/5-羟基阿魏酸-O-甲基转移酶;4CL—香豆酸辅酶A连接酶;CCOAOMT—咖啡酰辅酶AO-甲基转移酶;CAD—肉桂酸脱氢酶
COMT在木质素合成特异途径中主要是催化S木质素的生物合成。许多研究结果表明利用反义RNA技术抑制COMT和CCoAOMT基因的表达可以调整木质素的结构,降低转基因植株的木质素含量,并且这种木质素含量的适当减少不会影响植物正常的生长和发育。我国学者魏建华(2001)和赵燕华等(2004)先后将CCoAMOT反义基因导入毛白杨,转基因毛白杨内源CCoAOMT基因的表达在转录和翻译水平上均受到抑制,Klason木质素含量均有不同程度的降低。其中木质素含量下降幅度较大的株系,伴随着茎秆局部区明显发红,但对植物的生长和发育及其内部机械输导系统未产生明显的负效应。这些都说CCoAOMT的确能有效地调节植物中木质素的生物合成。
CAD参与木质素单体合成的最后一步是还原反应,当CAD基因突变时,CAD活性降低,木质素含量减少,G和S单体及松柏醛和芥子醛含量也明显下降。Baucher等(1996)从毛果杨X美洲黑杨分离出来的反义CAD转基因转入杨树(欧洲杨和银白杨),结果表明木质组织CAD活性减少70%,木质部是红色,对木质素的结构发生了某些改变而降低纸浆的能耗和生产成本。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。