



三、古生菌
古生菌(archaea)是由一群具有极端嗜热、嗜盐、嗜气特性微生物组成,它是一个系统发育上特殊分类单位,是根据16SrDNA片段的碱基序列(AAACUUAAAG)分析有别于细菌(AAACHCAAA),却与真菌的18SrRNA一致而划分出来的,可视为细菌类的一个分支。这是由R.HWhittaker等首先提出的三原界学说(图8-6),即在生物过程的早期,存在一类各种生物的共同祖先,由它分3条进化路线,形成3个原界。最初先分成两支,一支为真细菌原界;另一支为是古生菌—真核生物界,它在进行过程中又分为古生菌界(发展为今天的古生菌)和真核生物原界(包括真菌)。
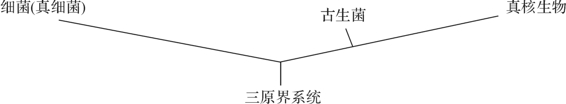
图8-6 三原界系统树
1.超嗜热古生菌
大多数超嗜热古生菌分离自地热区的土壤或含有元素硫和硫化物的水,并且大多数种可利用某种途径代谢硫。在陆地环境中,富硫泉、泥沼和土壤的温度可能会超过100℃,并且来自H2 S和S0的生物氧化作用产生H2 SO4会使环境变酸。这种热而且高硫的环境被称为硫黄温泉(salfataras)。一种嗜热水生菌(Thermus aquatics)的耐热DNA多聚酶“Taq”已被提出来,并在PCR扩增技术中得到应用。
超嗜热古生菌是专性厌氧。它们的产能代谢是化能有机营养型或化能无机营养型,以硫作为电子受体,进行厌氧呼吸或者电子供体进行化能无机营养代谢。元素硫被还原成H2S所需要的电子是从有机物的氧化或从H2获得。下列为几种超嗜热古生菌的产能反应:

硫化叶菌属(Sul folobus)和酸菌属(Acidianus)的生长温度从65℃开始最高达95℃,它们DNA中GC碱基比率低,分别为38%和31%。许多超嗜热细菌的DNA的GC碱基含量同样低,这是耐高温生物特性表现吗?某些溶质的积累对超嗜热细菌中蛋白质的稳定性有所帮助,如环式2,3-二磷酸甘油酸和伴侣蛋白质。据观察超嗜热古生菌的生存高温极端可达110℃。
2.极端嗜盐古生菌
高盐环境总是遍及世界各地的,极端嗜盐菌的最适生长则需要2~4mol/L(12%~23%)NaCl环境中缓慢生长。所以极端嗜盐菌不仅嗜盐,而且对盐的需求很高。所以,海盐场也是极端嗜盐原核生物的栖息地,它们也出现在腊肠、腌肉和咸鱼上。(https://www.xing528.com)
根据20世纪70年代16SrRNA序列和其他研究将极端嗜盐菌分为8个属,即盐杆菌属、盐球菌属、富盐菌属、盐盒菌属、盐深红菌属和嗜盐杆菌属等。盐杆菌属(halobacterinm)是最先被确认有代表性的属。盐杆细菌的细胞壁缺少肽聚糖,但含有醚键连接的类脂以及古生菌类型的RNA聚合酶,并对抗生素不敏感。所有的嗜盐古生菌均为革兰氏染色阴性、二分裂繁殖、无休眠状态,不产孢子。盐杆菌和盐球菌的基因组成很独特,在细胞DNA总量中含有25%~30%的频繁存在的大质粒,并且这些大质粒的GC碱基比率(57%~60%)与染色体DNAGC的比率(66%~68%)有明显差异。
所有的极端嗜盐古生菌均为化能有机营养。大多数为专性好氧,利用氨基酸、有机酸作为能源而少数种能氧化碳水化合物。嗜盐杆菌的电子传递链包括Cyta、b和c,在好氧生长的能量贮存是通过内膜化学渗透所引发的质子动力来进行。某些菌的厌氧生长是通过耗糖发酵以及通过硝酸盐的无氧呼吸链来进行的。极端嗜盐菌的生长需要大量的钠,若环境中用K+替代就不能满足对Na+的需求。K+进入细胞作为生理调节和保持高渗透压是需要的,但是低钠含量会使细胞壁破碎。嗜盐杆菌的细胞壁像所有古生菌一样不含有肽聚糖,以糖蛋白代替之。这种蛋白质含有高量的酸性氨基酸——天门冬氨酸和谷氨酸,带有负电荷成Na+屏蔽,结合在胞壁外表面以利于吸收。
3.产甲烷古生菌
通过16SrRNA的序列分析,已经得出产甲烷细菌的7个主要类群,如甲烷杆菌属(Methanobacterium)、甲烷嗜热菌属(Methanothermus)、甲烷球菌属(Methanococcus)和甲烷嗜盐菌属(Methanohalophilus)等。尽管产甲烷菌(methanogens)是专性厌氧生活并进行特殊的代谢方式,但它们在地球上还是普遍存在的,如木本、草本沼泽或瘤胃中,其中由反刍动物的暖气和湿地释放的甲烷是最大量的生物甲烷来源。在海水的沼积物中因含有相当多的硫酸盐细菌和硫酸盐还原细菌,它们与产甲烷的种群竞争可利用的乙酸盐和H2,因此,产甲烷菌不甚活跃。
现在证明有10种底物可作为一种甲烷细菌转化成甲烷,CO2几乎是甲烷细菌最普遍的底物,所需要的电子通常来自于H2,当有H2和CO2时,甲烷细菌进行自养型生长,以CO2作为碳源和电子受体。此外,甲基类底物,如甲醇、甲基胺和乙酸型底物乙酸等都可作为甲烷古生菌转化成甲烷的营养源,并释放出ATP能量。三类形式如下:
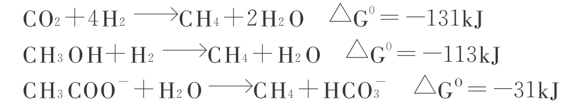
一般来说,从CO2还原成CH4是由分子氢来驱动,通过甲基团和甲基辅酶作用及乙酰辅酶A途径来完成,因为乙酰辅酶途径和产甲烷途径两者都会导致CH3基团的产生。两种途径的结合或许对微生物来说是一种能量的节约,但在制取乙酸的甲基团的生物步骤中,附加酶的合成是必需的。辅酶M(CoM)是甲基团的携带者,通过F430甲基还原复合物(CH3-S-CoM)最后将甲基还原成甲烷,并在最后一步进行能量贮存。
4.古生菌:地球早期的生命形式
现在知道,某些细胞大分子可作为进化时钟,衡量进化的变化。由于蛋白质合成过程所具有的古老特征以及其他一些原因,表明核糖体RNAs是鉴别生物间进化关系的重要分子。自20世纪70~80年代,分子系统学用核糖体亚单位的16SrRNA(原核生物)和18SrRNA(真核生物)序列作比较,经计算机分析,C.R.Woese(1977)提出了有别于传统生物进化系统三大类群(见本章第五节说明与图)。然而,L.Margulis等人(1970)提出真核细胞来自原核细胞的“内共学说(endosymbiotic theory)。”该学说认为,原始的厌氧原核生物以吞食其他原核生物为生,有时它们能容忍所捕获的原核生物在体内生活下去。共同生活的结果,被吞食的原核生物变成了细胞器,这就出现了真核细胞。内共生的主要依据是现代真核细胞的线粒体和叶绿体都很古老而且具有独立的自主活动,它们的DNA为环状,核糖体为70S,这些都是和细菌、蓝藻相同的。目前,以上两种观点还难以统一。
古生菌和真细菌之间的差别,不存在真细菌由古细菌进化而来。据G.E.Fox等(1980)推算它们之间16SrRNA的差异量,其起源发生可追溯到30多亿年以前。这三者各自进化,但古生菌与真核生物关系比真细菌间的关系更密切。表达了生命起源的复杂的多元途径。在这三个领域的生物中,现今的古生菌仍是最原始的,进化最低的。而真核生物已远离祖先,这种进化实为多细胞产生有关,可容纳大量的DNA。古生菌的生活环境是极端的,如高温、低pH和高盐,这是一种反应生命起源的环境条件,也是地球早期生命形式的最好进化遗迹。近几年,我国科学家在钻探工程中惊奇地发现在地下1080m和2000m深处的极端条件下,也发现了古生菌,有嗜酸、嗜铁、嗜甲烷,在缺氧高温的岩石细缝里,只靠吃铁、镍、甲烷等生存。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。