



二、光反应与光系统
光合作用的过程可分为3大步骤:原初反应(光能的吸收、传递和转换过程);电子传递和光合磷酸化(电能转化为活跃的化学能过程);碳同化(活跃的化学能转变为稳定的化学能过程)。第一、二个大步骤基本属于光反应,第三个大步骤属于暗反应,为酶催化的碳循环来完成,包括C3途径、C4途径和CAM途径。
1.光合作用的原初反应
原初反应(primary reaction)是光合作用的序幕,它包括光能的吸收传递和光化学反应。
(1)光能吸收
光由光粒子组成,有能量,故又称光量子。根据光子的能量公式:q=hv=hc/λ计算表明,光量子的能量与其波长成反比,因此,波长较短的蓝紫光比波长较长的红光能量高。叶绿素吸收光能受到激发后,除以热能、光能形式放出能量外,还把能量传给其他分子,也可捕获光能,发生光化学反应。光合色素有捕光色素复合体和反应中心色素复合体两类。捕光色素复合体由一群叶绿素a、b和胡萝卜素与色素蛋白结合而成,只起着吸收和传递光能的作用,这是光合原初反应的第一步。
捕光色素吸收的光能如何传到反应中心色素?不同的Chla、Chlb和类胡萝卜分子之间存在着非常复杂的激子相互作用,其传递迅速有效,传能时间在几百飞秒(fs)到1ps之间。Chen(1997)等采用飞秒激光技术对PSⅡ原初反应的超快动力学特性进行了研究,观察到一个760fs的组分,推测它可能属于天线色素分子向反应中心的能量传递过程。所以,大多数人认为激发能在完整的PSⅡ中,传递途径是由核心天线CP43、CP47接受捕光天线LHCⅡ和辅助外周天线CP29、CP26、CP24传来的激发能,并将激发能传递到反应中复合物。
(2)光化学反应
光化学反应是反应中心色素分子受光激发传递引起的氧化还原反应。由于光合作用光能转化的原初过程是由两个光系统反应以串联方式协同进行,其光能转化的效率取决于色素吸收的激发能在两个光系统之间的分配状况,通常当吸收的激发能在两个系统之间的分配处于平衡时,其光能转化的效率最高。
关于原初反应的反应中心的色素组成分,必须包含原始电子受体(A)和电子供体(D)。当反应中心色素分子(P)接受光能以后,首先变成激发态P*,并直接把电子传给原初电子受体A,使其还原为A-,而中心色素P+立即从原初电子供体夺取电子,使原初电子供体氧化D+而自身还原成P。那么上述原初光反应过程就可以用下面的反应通式来表示:
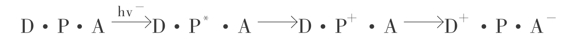
这样,只要有还原态的原始电子供体和氧化态的原初电子受体,反应中心就可以不断地进行光能转变,把光能转化为电能,随之通过电子传递体,传给最终电子受体NADP+,由此推动能量转换成生物能。其中,H2O是最终电子供体,D+是强氧化剂,它参与H2O的光氧化,释放出O2。
为了衡量光量子转化效率,必须确认光合单位。光合单位概念最早由Emergon等(1934)提出的。实践证明,同化1分子的CO2的一个完整单位是叶绿体,它约含2500~3000个叶绿素分子。或者从测定叶绿体的光合放O2发现,每释放一个O2分子需要8个光子,约2500个叶绿素分子参加反应。若按爱因斯坦光化学定律:一个分子吸收一个光子,就能引起一个光化学反应。因此一个“反应中心”应含有2500÷8=300个叶绿素分子。然而,最近对高等植物PSⅡ“反应中心”提纯的结果表明,一个高纯化的反应中心仅含有4~6个chla分子、2个Pheo分子(脱镁叶绿素)和2个β-Car分子(类胡萝卜素)。
(3)激发能的荧光现象
在通常情况下,一个未被光活化的叶绿素分子处于它的最低能级状态,即基态。但当它吸收一个红光光子被激发以后,跃迁到激发单线态。处于这种高能状态的分子是极不稳定的,它将通过下面三种途径去激发,释放它的能量而回到稳定的基态。
一种是非辐射去激发能量以热的形式耗损;另一种是发射荧光和磷光。叶绿素发射的荧光,其寿命约为10-9s。每100个吸收了光的叶绿素分子中约有30个会发出荧光,在叶绿素溶剂提取试管中,肉眼都能看到。叶绿素发射的磷光寿命比荧光长得多,约为10-3~10-2s,但强度只有荧光的1%。叶绿素的荧光是由第一单线态回到基态时发出的;而叶绿素的磷光则由三线态回到基态时发出的,因此,磷光的能量最低。
第三种是进行光化学反应,这是用于光合作用的能量部分。以上三种过程是由同时发生和竞争的。抑制光化学反应可以使荧光发射增强。因此,荧光产率变化的测定已成为光能转换研究中一种重要的监测手段。荧光和光化学量子产量的同时减少是由于激发态的热耗散引起的。所有非光化学荧光粹灭对于PSⅡ活性控制有着重要作用,但机制还不清楚。
2.两个光系统
(1)Emerson效应和两个光系统
早在1943年Emerson等在研究小球藻量子效率(每吸收1个光量子所释放的氧分子数)时发现,小球藻光合作用最有效的光波长为650~700nm红光和400~460nm蓝紫光。如果仅用波长大于685nm光照小球藻时,其量子效率大为降低,被称为红降现象;1958年,他们又发现补以短波红光,则量子效率明显增加,这种现象称为双光增益效应或爱默生效应。由此提出光合作用存在两种光系统合作进行的观念。这就成为60年代大量开展的关于两光系统的理论和实验基础。这期间,许多著名光合作用学者对叶绿体基粒类囊体膜进行分离,通过大量的光谱学(差异光谱)方法和细胞色素的氧化还原的电位及电子传递途径等测试。到60年代中期提出了光合作用中心的PSⅠ和PSⅡ以及其间电子传递体的排列顺序。Hill和Bendell首先提出两光系统(PSⅠ和PSⅡ)之间的串联关系,称为Z图,后人也称为Hill-Bendell图式。
(2)光系统Ⅱ和光系统Ⅰ
自20世纪60年代提出两个光系统(PSⅡ和PSⅠ)之后,在分子水平上对类囊体膜上的色素蛋白复合体的结构和能量转换机制的研究逐步深入。近些年来,我国学者在国家“973”项目安排下组织力量集中在“光合作用原初光能转化原理与调控”方面也做了不少工作而取得进展(匡廷云,2003)。
根据新近资料表明PSⅡ由外周捕光(天线)色素蛋白复合体(LHCⅡ),内周捕光(天线)色素蛋白复合体(PC43和CP47等),反应中心色素蛋白复合体(PSⅡ-RC)和Mn簇合物和外周蛋白33KD、24KD和17KD组成(图6-3)。通常将去LHCⅡ的PSⅡ部分称为PSⅡ核心复合物,而分离、纯化的PSⅡ-RC仅由D1蛋白(32KD)、D2蛋白(34KD)、细胞色素b559(Ctyb559)的α及亚基和PSb1基因产物等5种蛋白质及其结合的chla、脱镁叶绿素(Pheo)和β胡萝卜素(β-Car)组成。
研究还证明一个PSⅡ-RC中,只有2个chla分子具有光敏化性质,它们组成特殊的“分子对”,在原初的化学反应中起着电子供体作用,而它的原初电子受体是Pheo。由于这一特殊的chla“分子对”有一个680nm光谱吸收高峰,故称其为P680。目前已公认,一个具有放O2活性的“放氧中心复合体”则由PSⅡ反应中心,PSⅡ内周天线CP43及CP47,以及33KD外周蛋白组成,分子量为250~300KD。(https://www.xing528.com)
PSⅠ与PSⅡ相似,PSⅠ也由捕光色素蛋白复合体(LHC-Ⅰ)和反应中心色素蛋白复合体(PSⅠ-RC)组成,但与放O2有关的Mn簇合物和外周蛋白PSⅠ-RC中组成特殊chla“分子对”,在原初光化学反应中起原初电子供体功能的是P700。因为它的氧化还原吸收示差光谱中,在700mn有最大吸收,故而得名。
(3)光合链与光合磷酸化
①光合链
光合链是由两个光系统和若干电子传递体按一定的氧化还原电位依次排列而成的体系(图6-7)。Z链起点处,H2O是电子供体,其终点是NADP+受体。在整个电子传递过程中,只有两处:P680→P680*和P700→P700*(激发态),是逆氧化还原梯度有光能推动的需能反应,而其余的电子传递过程都是顺着能量梯度自发进行的。这个光合链的主要生理功能,是通过水的光解将光能转化为电能进行电子传递,并通过光合磷酸化产生ATP而最后将电子与质子传给NADP+产生NADPH,即所谓同化能力,用于暗反应的CO2固定。
D1蛋白和D2蛋白是PSⅡ的基本组成,P680(叶绿素a分子二聚体,dimer)就位于这两种蛋白之上。围绕P680的LCHⅡ吸收光能,激发P680为P680*,电子向类囊体膜外侧方向转移,传给pheo,pheo再把电子传给一种特殊的质体醌QA,QA进一步被还原为另一种质体醌QB。QB与来自基质的质子结合,形成还原质体醌(PQH2)。当PQH2转变为PQ时,会释放H+到腔内,有利于ATP合成。

图6-7 光合链Z型图
Cytbf是细胞色素b6f蛋白复合体,成分比较复杂,它由4个主要的大亚基:Cytf(33/34 KD)、Cytb6(23.5KD)、Rieske铁硫蛋白(Rieske Fe-S,20KD)和亚基IV(17KD);4个小亚基(MW<5KD):PetG、PetL、PetM和PetN;2~3个chla分子和1~3类胡萝卜素分子组成。此外,ATP合成酶由CF1和CF0两部分组成,参与电子传递过程的氧化磷酸化作用(图6-3)。我们回过头来看,当P680在射出电子以后,迅速从它的电子供体获取电子而复原。P680的直接电子供体目前尚不清楚,推测为D1蛋白中的酪氨酸残基(Try161)。在图6-7中以Z代表。Z供出电子以后被化成Z+。Z+是一个强氧化剂,它从H2O中拉电子使H2O氧化释放电子、质子和O2。
在PSⅠ中,“反应中心色素”分子P700受光激发后射出的高能电子,首先被一个距离最近的(~2nm)、称为A0的chla分子接受,然后将它传递给次级电子受体A1(叶醌),最后这个电子经过4Fe-4S原子簇(FX、FA和FB)传至Fd,并在Fd-NADP还原酶(FNR)催化下还原成NADPH。P700射出电子后引起的电子亏缺,将由它的次级电子供体PC提供的电子加以补充,使其恢复到原来的状态。PC供出电子以后被氧化成PC+。PC+是一种弱氧化剂(含铜原子的蛋白质),它从Cytf获得电子而使Cytf氧化。Fd也可把电子交给Cytb6而传回到PQ,形成围绕PSⅠ的循环电子传递。
②光合磷酸化
光合磷酸化由于电子传递方式不同,可分为非环式和环式磷酸化两种。这两个光合磷酸化电子传递的组成成分与排列,有过多次修改资料。据目前所知,在高等植物PSⅡ中,“反应中心色素”分子P680受光激发后射出的高能电子,首先被它的电子受体Pheo接受,形成Pheo-。Pheo-将电子传给QA,再传给PQ,并产生非环式磷酸化ATP。PQ是质体醌A和C的混合物,每条光合链大约含6个这样的PQ分子组成PQ库。PQ的光还原氧化作用促使膜内外的质子进行频繁的交换,形成膜内外质子梯度。然而,PQ的氧化还原过程是由连接两个光系统中间电子传递体Cyt6f催化的;随后,把电子传给PC(质蓝素),PC是PSⅠ的原初电子供体,把电子传给P700,使PSⅡ与PSⅠ得到连接。在这个过程中,电子传递是一个开放的通路,故称为非循环光合磷酸化(noncyclic photophosphorylation)。非循环光合磷酸化在基粒片层进行,它在光合磷酸化中占主要地位。另外,PSⅠ产生的电子经过一些传递体传递后,伴随形成腔内外H+浓度差,只引起ATP的形成,而不放O2,也无NADP+还原反应。在这个过程中,电子经过一系列传递后降低了能位,最后经过PC重新回到原来的起点,也就是电子的传递是一个闭合的回路,故称为循环光合磷酸化(cyclic photophosphorylation)。循环光合磷酸化在基质片层内进行,在高等植物中可能起着补充ATP不足的作用。
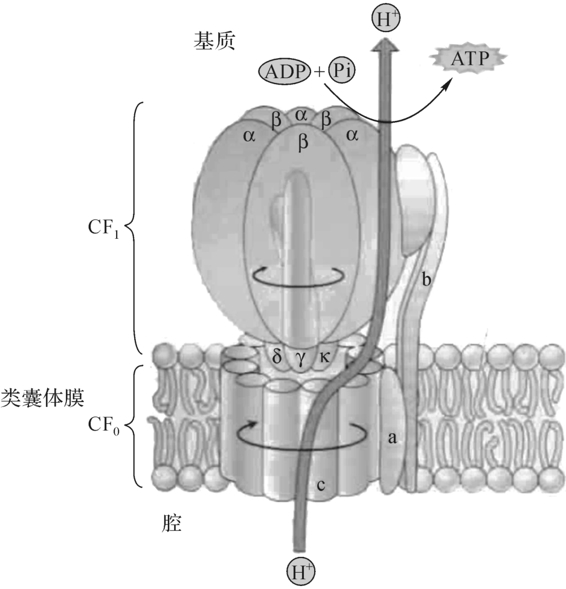
图6-8 ATP合酶的结构
ATP合酶是一个大的多亚单位的复合物,它的功能是把ADP和Pi合成为ATP,故名ATP合酶(ATPsynthase)。它也将ATP的合成与电子传递和H+跨膜转运偶联起来,故又称为耦联因子(coupling factor)。ATP合酶复合体由头部(CF1)和柄部(CF0)组成(图6-8)。其中CF1在类囊体的表面,CF0伸入类囊体内,CF1是由5种多肽(α、β、γ、δ、ε)组成,它们的数目比为3αЗβγδε。CF0可能由4种多肽(a、b、b′、c12)组成,CF0由几个多肽组成横跨类囊体的通道,以通过质子。组成通道的多肽包括3份α和β多肽,随意排列,似桔子一样。催化的功能大多数是由β多肽执行,其他多肽则起调节作用。
线粒体ATP合酶的分子结构已经用X-线晶体学研究查明,虽然叶绿体和线粒体之间的酶有很大的区别,但两者的结构是相同的,它们在电子传递伴随着质子传递也是一致的。还有一点相似的是ATP合酶的作用机理,即在催化ATP合成时,柄的内部和大部分CF0的酶都在转动着,这些酶实质上是微小的分子发动机。
关于光合磷酸化的机理,可用英国人P.Mitchell(1961)提出的化学渗透假说(chemiosmotic hypothesis)来解释。在类囊体的电子传递体中,PQ可传递电子和质子,而其他传递体,如PC和Fd等,只传递电子而不传递质子。光照引起水的裂解,水释放的质子留在膜内侧,水释放的电子进入电子传递链中的PQ。PQ在接受水裂解传来的电子的同时,又接受膜外侧传来的质子。PQ将质子排入膜内侧,将电子传给PC。这样,膜内侧质子浓度高而膜外侧低,膜内侧电位较膜外侧高。于是膜内外产生质子浓度差(ΔpH)和电位差(Δψ),两者合称为质子动力(proton motive force,PMF),即为光合磷酸化的动力。当H+沿着浓度梯度返回膜外侧时,在ATP合酶催化下,ADP和Pi脱水形成ATP。
经过上述变化以后,由光能转变来的电能便进一步形成活跃的化学能,暂时贮存在ATP和NADPH中。由于ATP和NADPH用于暗反应中化CO2的同化,所以,把这两种物质合称为同化能力(assimilatory power)。
(4)平衡两个光系统的激发能的分配与生理学意义
一般植物光合机构的LHCⅡ磷酸化主要受电子递体质醌(PQ)和细胞色素b6f复合体氧化还原状态的调节,从而影响其在两种光系统间的移动。近些年来,人们发现植物光合机构的状态转换也可以通过两光系统相互接近导致激发能满溢来平衡两个光系统的激发能分配(Wollman FA2001)。譬如,当外界离子浓度骤变可以引起盐藻LHCH磷酸化,其调节过程与电子递体的氧化还原状态无关;绿藻的状态转换可以调节细胞内的ATP供求关系(刘贤德2006)。
在自然环境中,植物光合作用总是在接受不断变化的日照条件下进行的。由于两个光系统结合色素组成不同,这种光强与光质的不断变化常常导致两个光系统的不均衡激发。所以,植物光合机构的状态转换作为一种反馈调节机制才能平衡激发能在两个光系统间的分配。当PSⅡ被过度激发时,从PSⅡ传来的电子量超过从PSⅠ传出的电子量,从而导致中间电子递体PQ与Cytbf复合体处于还原状态,LHCⅡ被磷酸化,光合机构向状态转换。磷酸化的LHCⅡ从PSⅡ脱离并转移到PSⅠ上,成为PSⅠ的捕光天线;经过这样变动,PSⅡ的光吸收截面降低,而PSⅠ的光吸收截面增加,结果本来分配给PSⅡ的一部分光能被传递给PSⅠ,从而相对增加了PSⅠ的光吸收。
与上述相反,当PSⅠ被过度激发时,细胞间状态PSⅠ转换,电子递体PQ及Cytbf被氧化,LHCⅡ激酶没有活性,磷酸化的CHCⅡ在磷酸酯酶的作用下去磷酸化,并重新与PSⅡ结合,从而相对增加PSⅡ的光吸收截面及光吸收。在自然光质光量不断发生变化的环境中,植物用这种方式可以调节激发能在两个光系统间的分配,从而较好地利用光能。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。