



一、基因工程酶学
1.限制酶和DNA连接酶
(1)限制性核酸内切酶
限制性核酸内切酶简称限制酶(restriction enzyme),是一类能够识别双链DNA分子中的某种特定核苷酸序列,并在相关位置切割DNA双链结构的核酸内切酶。它们主要是从原核生物中分离纯化获得的,至今已从近300种不同微生物中分离出了2300种限制性核酸内切酶。在限制酶的作用下,侵入细菌的外源DNA分子被切割成大小不同的片段,使其基因失去功能,而细菌本身的DNA因受到一种甲基化酶的保护而免遭降解。限制酶的发现(1968)和应用使得DNA分子的体外切割成为可能,也使体外重组技术的应用成为可能。
目前根据酶的识别序列及切割位点、催化条件及是否具有修饰酶活性,将限制酶分为Ⅰ、Ⅱ、Ⅲ型。Ⅱ型限制酶即通常所说的限制酶,具有这类酶的微生物其限制、修饰系统分别由两种不同方式完成,即限制性内切酶和修饰性甲基化酶。Ⅱ型限制酶的特点是相对分子质量小,仅需Mg2+作为催化反应的辅助因子,能识别并切割DNA链上的特异性核苷酸顺序,产生特异性DNA片段,因此,它在基因工程中有着广泛的应用。
多数Ⅱ类酶的识别序列为4、5或6个碱基对,并且具有180°旋转对称的回文结构,即有一个中心对轴,序列正读和反读是一样的。Ⅱ型限制酶对DNA的切割有两种方式:
①黏性末端:限制酶在识别序列时进行交错切割,结果形成两条单链末端,这种末端的核苷酸顺序是互补的,所以称为黏性末端,如ECORI的识别顺序为:

垂线表示中心对称轴,两链读核苷酸从5′到3′顺序都是GAATTC,这就是回文顺序。在双链交错切割生成5′—G和AATTC—3′、3′—CTTAA和G—5′两个DNA片段,各有一个单链末端,二条单链是互补的,可通过DNA连接酶的作用而黏合。
②平端:Ⅱ型酶切割方式的另一种是同一位置切割双链,产生平端,例如EcoRV的识别位置是:
5′—GAT′|ATC—3′
3′—CTA′|TAG—5′
切割后形成5′—GAT和ATC—3′、3′—CTA和TAG—5′。这种平端同样可以通过DNA连接酶连接起来。
(2)DNA连接酶
DNA连接酶广泛存在于各种生物体内,其催化的基本反应是将DNA双链上相邻的3′—羟基和5′—磷酸基因共价结合成了3′、5′—磷酸二酯键,使原来断开的DNA缺口重新连接起来,因此,它在DNA复制、修复以及体内外重组过程中起着重要作用。
最常用的DNA连接酶是T4DNA连接酶和大肠杆菌DNA连接酶。T4DNA连接酶是由大肠杆菌(E·coil)T4噬菌体DNA30编码的产物,最早从T4噬菌体感染的大肠杆菌中分离纯化的,相对分子质量为60KD,是一种单链多肽酶能催化两个DNA片段的3′—OH末端和5′—P末端形成磷酸二酯键,需要ATP作为辅助因子并由ATP提供能量。T4噬菌体DNA30现已被克隆,并可在大肠杆菌中大量表达,因此T4DNA连接酶比较容易制备。此酶可高效催化dsDNA片段间黏末端的连接,并可催化平端DNA片段的连接。
大肠杆菌DNA连接酶又称为E.coliDNA连接酶,是由大肠杆菌染色体基因组中Lig基因编码的,相对分子质量为75KD。Lig基因已被克隆,并能在相应大肠杆菌细胞中大量表达。此酶不能催化平端DNA分子之间的连接,其底物只能是带缺口的dsDNA分子和具有同源互补黏末端的不同DNA分子,并以NAD+作为辅助因子。
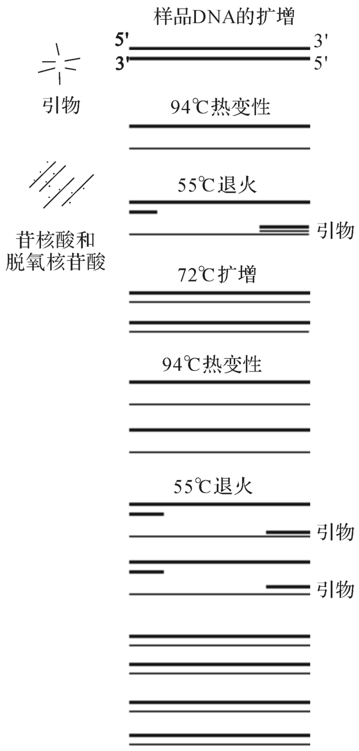
图4-8 PCR反应过程
在真核生物细胞中也存在DNA连接酶,且有两种类型,分别称为连接酶Ⅰ和Ⅱ,反应中利用ATP提供能量。DNA连接酶Ⅰ分子质量约为200KD,主要存在于生长旺盛细胞中,DNA连接酶Ⅱ分子质量约85KD,主要存在于生长不活跃的细胞中。它们的催化反应在DNA复制和修复过程中都起重要作用。
2.DNA聚合酶和反转录酶
(1)DNA聚合酶
DNA聚合酶(DNApolymerase)催化以DNA为模板合成DNA反应,在分子克隆技术中主要用于DNA的体外合成、探针的标记等。经常使用的DNA聚合酶有大肠杆菌DNA聚合酶Ⅰ(全酶)、大肠杆菌DNA聚合酶的klenow片段(即klenow酶)、T4DNA聚合酶、耐热DNA聚合物以及反转录酶等。这些DNA聚合酶反应的共同特点是:要以四种脱氧核糖核苷酸作底物;聚合反应需要DNA模板的指导;要有引物3′—羟基的存在;新链的合成方向为5′→3′。由此看出DNA聚合酶合成的产物是模板性质相同的复制品。(https://www.xing528.com)
大肠杆菌DNA聚合酶Ⅰ,简称DNAp01Ⅰ,又称kornberg酶,它是大肠杆菌P01 A基因编码,由一条多肽链组成,分子质量为109KD。在大肠杆菌中,每个细胞约有400个分子的DNA聚合酶Ⅰ。在37℃下,1分子DNA聚合酶1分钟可以催化约1000个核苷酸的聚合。DNA聚合酶Ⅰ是一个多功能酶,具有三种活性,即5′→3′聚合、5′→3′外切活性和3′→5′外切活性。
大肠杆菌聚合酶Ⅰ全酶经枯草杆菌蛋白酶处理,可以产生分子质量为67KD和35KD两个大小不同的片段。其中大片段由Klenow等人于1970年报道,故称为Klenow片段,也称Klenow聚合物。该酶具有两种催化活性:①5′→3′聚合酶活性,可以合成DNA;②3′→5′外切酶活性,可从3′端降解DNA,但是,它失去了5′→3′外切酶活性。此外,该酶在DNA克隆中的主要用途:通过聚合作用填补或标记DNA的3′隐蔽末端;催化合成cDNA的第二链及DNA序列测定。
大肠杆菌DNA聚合酶Ⅱ(DNApolⅡ):DNA聚合酶Ⅰ缺陷的突变株仍能生存,这表明DNApolⅠ不是DNA复制的主要聚合酶。人们开始寻找另外的DNA聚合酶,并于1970年发现了DNApolⅡ。此酶分子质量为120KD,每个细胞内约有100个酶分子,但活性只有DNApolⅠ的5%。该酶的催化特性如下:
①聚合作用:该酶催化DNA的聚合,但是对模板有特殊的要求。该酶的最适模板是双链DNA中间有空隙(gap)的单链DNA部分,而且该单链空隙部分不长于100个核苷酸。对于较长的单链DNA模板区该酶的聚合活性很低。但是用单链结合蛋白(SSBP)可以提高其聚合速率,可达原来的50~100倍。
②该酶也具有3′→5′外切酶活性,但无5′→3′外切酶活性。
③该酶对作用底物的选择性较强,一般只能将2-脱氧核苷酸掺入到DNA链中。
④该酶不是复制的主要聚合酶,因为此酶缺陷的大肠杆菌突变株的DNA复制都正常。可能在DNA的损伤修复中该酶能起到一定的作用。
大肠杆菌DNA聚合酶Ⅲ(DNApolⅢ):DNApolⅢ全酶由多种亚基组成,而且容易分解。大肠杆菌每个细胞中只有10~20个酶分子,因此不易获得纯品,给研究该酶的各种性质和功能带来了许多困难。直到不久前,才对其性质和功能有所了解,但每个亚基的具体作用仍不十分清楚。尽管该酶在细胞内存在的数量较少,但催化脱氧核苷酸掺入DNA链的速率分别是DNA聚合酶Ⅱ的15倍和30倍。该酶对模板的要求与DNA聚合酶Ⅱ相同,最适模板也是链DNA中间有空隙的单链DNA,单链结合蛋白可以提高该酶催化单链DNA模板的DNA聚合作用。DNApolⅢ也有3′→5′和5′→3′外切酶活性,但是3′→5′外切酶活性的最适底物是单链DNA,只产生5′—单核苷酸,不会产生二核苷酸,即每次只能从3′端开始切除一个核苷酸。5′→3′外切酶活性也要求有单链DNA为起始作用底物,但一旦开始后,便可作用于双链区。DNA聚合酶Ⅲ是细胞内DNA复制所必需的酶,缺乏该酶的温度突变株在限制温度(non permissive temperature)内是不能生长的,此种突变株的裂解液也不能合DNA,但加入DNA聚合酶Ⅲ则可以恢复其合成DNA的能力。
真核生物的DNA聚合酶:真核生物中也具有几种DNA聚合酶,但这些聚合酶都没有3′→5′或5′→3′外切酶活性。其聚合反应机制与原核生物的聚合一样。主要的酶(占总量的80%~90%)是DNA聚合酶α,分子量为300KD,含有4个或5个亚基,主要负责染色体DNA的复制。DNA聚合酶β,分子量为45KD,仅含有一条链,主要作用是修复核内DNA。第三种聚合酶γ,分子量为140KD,存在于线粒体内,负责催化线粒体DNA的复制。最近从兔的骨髓细胞质中分离到一种新的DNA聚合酶,这种酶与上述真核生物的DNA聚合酶不同,而与原核生物的聚合酶类似,具有3′→5′核酸外切酶活性,称为DNA聚合酶δ。另外,也从酵母、原虫等低真核生物中分离到一些DNA聚合酶。一般来说,这些低等生物都没有低分子量的DNA聚合酶β,而且与原核生物的DNA聚合酶相似,有3′→5′外切酶活性。
(2)Taq DNA聚合酶
Taq DNA聚合酶是第一个被发现的热稳定DNA聚合酶,分子质量为65KD,1986年由Erlish等从温泉中分离的一株水生噬热杆菌(Thermus aquaticus)中提取获得。此酶具有5′→3′聚合活性和5′→3′外切活性,且能耐高温,在70℃反应2h后其残留活性还能保持原来的90%以上。1998年Saiki等将Taq DNA聚合酶成功地用于PCR技术,对DNA的特定序列进行体外扩增。在PCR循环过程中,Taq DNA聚合酶在变性步骤中(约94℃)不易失活,可直接进入下一轮循环,因此不必每轮循环时重加入新酶。这使得Taq DNA聚合酶成为PCR技术中的独特用酶(图4-8)。
(3)反转录酶
反转录酶(reverse transcriptase),又称为依赖RNA的DNA聚合酶。1970年Tem in等人在致癌RNA病毒中发现的一种特殊的DNA聚合酶,它以RNA为模板,dNTP为底物,tRNA为引物(主要是色氨酸tRNA),在tRND3′—OH末端上,根据碱基配对原则,按5′→3′方向,合成一条与RNA模板互补的单链,这条DNA单链叫互补DNA即cDNA。随后,在反转录酶的作用下,水解掉RNA链,再以cDNA为模板合成第二条DNA链,如图4-9所示。这一过程与一般遗传信息流转录(由DNA→RNA)的方向相反,因此,由RNA→DNA的过程称之为反转录作用。现已发现反转录酶不仅普遍存在于病毒之中,而且哺乳动物的胚胎细胞和正在分裂的淋巴细胞中也有反转录酶。
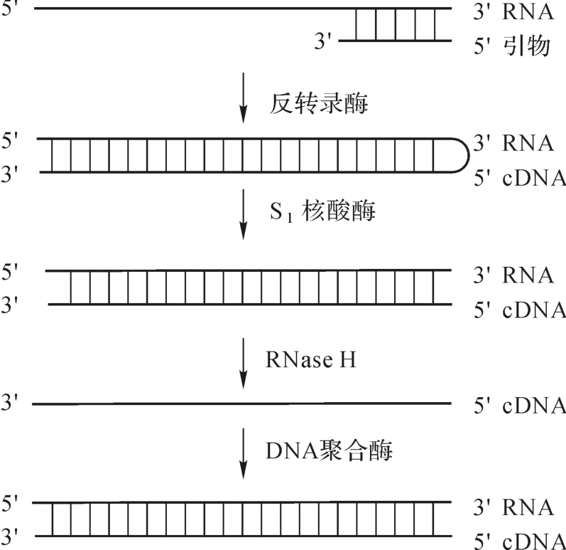
图4-9 反转酶催化DNA分子合成
基因工程中有两种商品化的反转录酶:一种来自纯化的禽成髓细胞瘤病毒(avian myoblastosis virus,AMV)提取的;另一种是Moloney鼠白血病病毒(moloney murine leukemia virus,Mo-MIV)克隆的反转录酶编码基因在大肠杆菌的表达产物。这两种反转录酶的主要区别:禽源反转录酶在42℃(鸡的正常体温)能有效地发挥作用,而鼠源的则迅速失活。携带反转录酶的病毒又称反转录病毒,它侵入宿主细胞后,先以病毒RNA为模板依靠反转录酶催化合成DNA,随后这种DNA环化并整合到宿主细胞的染色体DNA中去,以原病毒的形式在宿主细胞中一代代传递下去。近来发现许多反转录病毒基因组中都含有癌基因(oncogene),如果由于某种因素激活了癌基因就可使宿主细胞转化为癌细胞。
3.修饰酶类
在基因克隆技术中,除了限制酶、连接酶、聚合酶等主要工具酶外,还经常使用某些酶的相关功能对DNA和RNA分子进行修饰,故有修饰酶类之统称。例如,以末端转移酶的功能为基础建立了同聚物着尾法连接DNA片段,使用碱性磷酸酶切除DNA片段5′—端磷酸基以有效地防止载体DNA的自身环化等。
(1)S1核酸酶是由米霉霉(Aspergillusoryzae)中提取的,它是一种特异性单链核苷酸外切酶,能降解DNA和单链RNA,产生5′单链核苷酸或寡核苷酸。S1核酸酶是在基因工程中用途很多,如消化限制酶产生的黏末端,使之成为平端,降解cDNA合成过程中形成的发卡环结构;在RNA—DNA杂交后,结合S1核酸酶水解,可确定内含子的位置。
(2)碱性磷酸酶(alralime phosphatase)有两种:一种是从大肠杆菌中分离的,叫细菌碱性磷酸酶(BAP),另一种是从小牛肠中分离的,叫小牛肠碱性磷酸酶(CAP)。其共同特点是,特异性切去DNA或RNA分子的5′—P,从而使DNA或RNA分子5′—P末端转换为5′—OH末端,即所谓核酸分子脱磷酸作用。碱性磷酸酶在基因工程中的应用主要在于制备5′—末端标记的核酸探针和防止DNA重组中载体的自身环化。
(3)T4多聚核苷酸激酶(T4poly nucleotide kinase)简称T4激酶,来源于T4噬菌体感染的大肠杆菌细胞,并且已在多种哺乳动物中发现。T4激酶可催化γ位上磷酸从ATP分子转移给DNA或RNA的5′—OH末端。这种作用不受底物分子链的长短限制,甚至单核苷酸也同样适用。该酶广泛应用于DNA和RNA的末端标记,标记时以[γ-32p]ATP作为底物。
(4)末端脱氧核苷酸转移酶,简称末端转移酶(termind transferase),是从小牛胸腺中纯化出的一种相对分子质量较小(34×103)的碱性蛋白酶。末端转移酶催化5′—脱氧核苷三磷酸转移到另一个DNA分子的3′—OH末端,即进行5′→3′方向聚合作用。与DNA聚合酶不同,它不需要模板存在,就可以催化DNA分子聚合。在基因工程中,利用末端转移酶不需要模板,但4种dNTP中任何一种都可作为反应前体物的特性,可以给平端DNA片段3′—OH加上同聚物PolyC或PolyG或PolyT、PolyA,形成同聚物尾巴,造成人工黏末端,有利于DNA重组。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。