



带有两捕食趋向的食物链捕食模型古典解的全局存在性
李成林 尹国成
摘 要:本文考察了在留曼边界条件下带有两捕食趋向和Holing-II型反应函数的三种群食物链模型。应用抛物方程Schauder估计及Lp估计,证明了此模型存在唯一全局古典解。
关键词:食物链 古典解 Schauder估计 Lp估计
一、引 言
在捕食过程中,食饵和捕食者除了自由扩散外,空间的临时变化速度取决于食饵的梯度,即食饵趋向( Prey-taxis)。通常假设它与食饵的梯度成正比。在生态现象中,捕食者被食饵吸引。一般来说,食物越丰富,对捕食者的吸引就越大。在文献[13]中,作者对种群运动个体行为的测定证实了这一假设。因此,具有食饵趋向的捕食模型被提出,它能很好地揭示捕食者捕食的行为过程。最近,一些学者、生态家、数学家研究了与生态物种相关的食饵趋向。食物链是种群生态学中常见的现象,也是生态学中研究群体结构的一个中心课题之一。我们将研究食物链中的一基本例型,即三物种食物链,但包括食饵趋向。
在文献[1]中,作者证明了带有一个食饵趋向的捕食—食饵模型弱解的存在唯一性。在文献[4]中,作者把文献[1]的结果推广到一个n×m反应扩散食饵趋向系统。在文献[14]中,作者证明了带一个食饵趋向的捕食—食饵两种群模型古典解的存在唯一性。在这篇文章中,我们考察如下带有两个食饵趋向和Holing-Ⅱ型反应函数的三物种食物链系统:
其中Ω是Rn中的有界区域,光滑边界为aΩ,u1和ui(i= 2,3)分别代表捕食者和食饵密度(u2既是食饵密度,也是捕食者密度),正常数a,Ki,ri,mi,ei,mi/ci,bi/ci,mi/bi(i=2,3)分别代表捕食者的死亡率,食饵的环境容纳量,食饵增长率,半饱和系数,转化率,一个捕食者抓捕一个食饵所用的时间,操作时间,完成半饱和率所需的食饵密度;捕食者被食饵所吸引,正常数βi(i= 1,2)记为食饵趋向的敏感性;流β1 u1▽u2和β2 u2▽u3取决于u2和u3种群增加的密度,按此方式,捕食者朝向食饵密度较高的地方进行捕食。
虽然在文献[14]中作者证明了带有一个食饵趋向的食饵—捕食两种群模型具有唯一的古典解,但只有一个食饵趋向且是两个种群。在这篇文章中我们所讨论的模型是具有两食饵趋向的三种群模型,种群维数增加,给证明带来了根本性的改变,为完成证明,我们采用文献[5,15]中的技能来证明。
在整篇文章中,我们假设:
当u1≥u1m时,β1= 0;当u2≥u2m时,β2=0. (1.2)
当ui≥uim时,βi=0(i= 1,2),这一假设具有确切的生态解释:当捕食者密度在Ω内某点达到临界值uim时,捕食者停止捕食,且当ui≥uim时食饵趋向系数βi为0。
在这篇文章中,我们也假设:
其中,0<α<1 。
。
这篇文章的主要结论如下:
定理1.1:假设(1.2)和(1.3)成立,则对AT>0,系统(1.1)存在唯一解U=(u1,u2,u3)∈C2+α,1+α(QT),且对Ax∈Ω及t>0有
u1(x,t)≥0,u2(x,t)≥0,0≤u3(x,t)≤K3.
H.Amann在文献[16,17,18]中已证明了包括系统(1.1)在内的解的局部存在性,但全局解的存在性仍未证明,因此本文只证明系统(1.1)的解的全局存在性。
二、全局解的存在性
首先我们对系统(1.1)建立先验估计。为方便起见,记与T有关的常数为N,与T无关的常数为N0。
引理2.1:假设U=(u1,u2,u3)∈C2,1(QT)是系统(1.1)的一个解,那么
u1(x,t)≥0,u2(x,t)≥0,0≤u3(x,t)≤K3.
证明:由系统(1.1)可得
显然,u1=0是(2.1)的一个下解,利用最大值原理,可得u1≥0,类似可得u2≥0和u3≥0。
另一方面,由系统(1.1)中的第三个方程可得
由(2.2)知K3是(2.2)的一个上解。因此有0≤u3(x,t)≤K3.
证毕。
引理2.2:假设U=(u1,u2,u3)∈C2,1(QT)是系统(1.1)的一个解,那么对A p>1,有
‖u1‖LP(QT)≤N,‖u2‖LP(QT)≤N,‖u3‖LP(QT)≤N.
证明:在系统(1.1)的第二个方程两边同乘u2p-1(p> 1),在QT上积分可得
当u2≥u2m时,有
因此有
用Gronwall引理,有
所以,对u2<u2m,有
类似地,有
显然,Ωu3(x,t)dxdt≤ΩK(x,t)dxdt≤N.
证毕。
引理2.3:假设U=(u1,u2,u3)∈C2,1(QT)是系统(1.1)的一个解,那么对A p>5,有
证明:系统(1.1)的第三个方程可写为 -d3Δu3-(r3-r3u3/K3-c3u3/(m3+b3u3)) u3=0.
-d3Δu3-(r3-r3u3/K3-c3u3/(m3+b3u3)) u3=0.
其中‖r3-r3 u3/K3-c3 u2/(m3+ b3 u3)‖Lp(QT)≤N,由抛物方程Lp估计,有
‖u3‖Wp2,1(QT)≤N.
应用Sobolev嵌入定理(取p>5),有
‖▽u3‖L∞(QT)≤N.
由系统(1.1)的第二个方程可得
 -d2Δu2+β2▽u2·▽u3
-d2Δu2+β2▽u2·▽u3
=-β2u2Δu2+ r2(1-u2/K2) u2-
(t,x)∈(0,T)×Ω,
 = 0
= 0
(t,x)∈(0,T)×aΩ,
u2(0,x)= u20(x)≥0
x∈Ω.
(2.3)(https://www.xing528.com)
其中, ‖Lp(QT)≤N.
‖-β2u2Δu3+ r2(1-u2/K2) u2-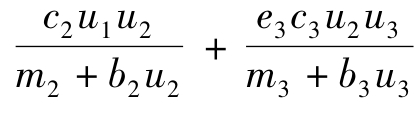
由抛物方程的Lp估计,有
‖u2‖Wp2,1(QT)≤N.
由Sobolev嵌入定理(取p>5),有
‖▽u2‖L∞(QT)≤N.
类似地,由(1.1)的第一个方程,得
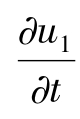 -d1Δu1+β1▽u1·▽u2=-β1u1Δu2-a+
-d1Δu1+β1▽u1·▽u2=-β1u1Δu2-a+
(t,x)∈(0,T)×Ω,
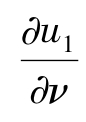 = 0
= 0
(t,x)∈(0,T)×aΩ,
u1(0,x)= u10(x)≥0
x∈Ω.
(2.4)
其中,
‖-β1u1Δu2-a+ ‖Lp(QT)≤N.
‖Lp(QT)≤N.
再由抛物方程的Lp估计可得
‖u1‖Wp2,1(QT)≤N.
证毕。
引理2.4:假设U=(u1,u2,u3)∈C2,1(QT)是系统(1.1)的一个解,那么对A p>5,有
‖u1‖C2+α,1+α/2(QT)≤N,‖u2‖C2+α,1+α/2(QT)≤N,‖u3‖C2+α,1+α/2(QT)≤N.
证明:由Sobolev嵌入定理(取p>5)和引理2.3,得
由(2.5)和系统(1.1)的第三个方程,用Schauder估计得
‖u3‖C2+α,1+α/2(QT)≤N.
对(2.3)用抛物方程Schauder估计,结合(2.5)可得
‖u2‖C2+α,1+α/2(QT)≤N.
由(2.5)和(2.4),用抛物方程用Schauder估计得
‖u1‖C2+α,1+α/2(QT)≤N.
证毕。
因此,我们可用[5,15]的方法把局部解延拓到A t≥0,即有定理2.1。
定理2.1:假设(1.2)和(1.3)成立,那么对A T>0,系统(1.1)存在唯一解U=(u1,u2,u3)∈C2+α,1+α/2(QT),且对A x∈Ω和A t>0,有
u1(x,t)≥0,u2(x,t)≥0,0≤u3(x,t)≤K3.
参考文献:
[1]B.E.Ainseba,M.Bendahmane,A.Noussair.A reaction-diffusion system modeling predator-prey with prey-taxis[J].Nonlinear Anal,RWA,2008(9).
[2]H.Amann.Fixed point and nonlinear eigenvalue problems in ordered Banach spaces[J].SIAM Rev,1976(18).
[3]A.Anderson,M.Chaplain,C.Garcia-Reimbert,C.Vargas.A gradient-driven mathematical model of antiangiogenesis[J].Math. Comput. Model,2000(32).
[4]M.Bendahmane.Analysis of a reaction-diffusion system modeling predator-prey with prey-taxis[J].Net. Hetero. Med,2008(3).
[5]A.Friedman,G.Lolas.Analysis of amathematical model of tumor lymphangiogenesis[J].Math. Models Methods appl. Sci,2005(1).
[6]T. Hillen,K.Painter.Global existence for a parabolic chemotaxismodel with prevention of overcrowding[J].Advin. Appl. Math,2001(26).
[7]C.S.Holling.Some characteristics of simple types of predation and parasitism[J].Canadian Entomologist,1959(91).
[8]D.Horstmann,M.Winkler.Boundedness vs. blow-up in a chemotaxis system[J].J.Differential Equationa,2005(215).
[9]E.F.Keller,L.A.Segel. Initiation of slime mold aggregation viewed as instability[J].J.Theoret.Biol,1970(26).
[10]W.Ko,K.Ryu. A qualitative study on geneal Gause-type predator-prey models with non-monotonic functional response[J].Nonlinear Anal,RWA,2009(10).
[11]O.A.Ladyzenskaja,V.A.Solonnikov,N.N.Ural'ceva.Linear and Quasi-Linear Equations of Paraboblic Type[M].in: Amer. Math. Soc. Transl.,Vol. 23,Providence,RI,1968.
[12]López-Gómez.Positive periodic solutions of Lotka-Volterra reaction-diffusion systems[J].Differential Integral Equations,1992,5(1).
[13]A.Okubo,H.C.Chiang.Analysis of the kinematics of Anarete pritchardi Kim[M]. col. 10,Springer,Berlin,1980.
[14]Y.Tao.Global existence of classical solution to a predator-prey model with nonlinear prey-taxis[J].Nonlinear Analysis: Real World Applications(2009),doi: 10.1016/j. nonrwa.. 05. 005.
[15]Y.Tao,M.Wang.Global solution for a chemotactic-haptotactic model of cancer invasion[J].Nonlinearity,2008(21).
[16]H. Amann.Dynamic theory of quasilinear parabolic equationsⅠ: Abstact evolution equations[J].Nonlinear Anal,1988,12(9).
[17]H.Amann.Dynamic theory of quasilinear parabolic equationsⅡ: Reacton-diffusion systems[J]. Differential Integal Equations,1900(3).
[18]H.Amann.Dynamic theory of quasilinear parabolic systems-Ⅲ[J]. Global existence,Math. Z.1989(202).
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。