



4.3.4 鼠科
鼠科(Muridae) 是啮齿目中第2个大科,多为中小型种类。尾一般较长,尾多裸露或被稀疏毛。环状排列的尾鳞外露明显,体形多修长,鼻吻部略尖突。耳壳较长,无颊囊。头骨较狭长,额部扁平或稍隆起,颧弓细弱,无眶上突,眶下孔上宽下窄略呈“V”字形。臼齿有齿根,无前臼齿,第1、第2上臼齿(M1、M2)咀嚼面有3纵列齿尖,每3个并列齿尖形成1个横嵴。
鼠科种类较多,适应能力强,分布广泛,繁殖潜力大。除少数种类营树栖、半树栖或两栖生活外,大多数种类均营陆栖穴居生活。栖息地十分广泛,森林、草原、半荒漠、农田、建筑物中均有其踪迹,常盗食粮食、污染食品,咬坏各种物资、家具等,不少种类还能携带和传播鼠疫、流行性出血热等自然疫源性疾病。所以本科的多数种类为重要的农林业和医学害鼠。
鼠科在全世界计有6亚科107属435种,国内计有1亚科(鼠亚科Muridae)12属43种,河南省有1亚科5属10种。
河南省鼠科分属检索表
1a.体型较大,成体体长超过150 mm,后足长超过25 mm,颅全长超过28 mm 3……

4.3.4.1 姬鼠属Apodemus
河南省姬鼠属分种检索表

(1)黑线姬鼠(Apodemus agrarius,Pallas 1771)
地方名:黑线鼠、姬鼠。
英文名:Striped field mouse
1)鉴别特征 身体背部中央有1条黑色纵行条纹,耳较小,尾长略短于体长,第3上臼齿(M3)咀嚼面有2齿叶。
2)形态描述(图4.53)

图4.53 黑线姬鼠
①外形 体型较小,体长75~120 mm,尾长略短于体长,约为体长的2/3,尾毛不发达,鳞片裸露,耳壳较小,前折不达眼部,四肢略细弱,后足较短,雌性具乳头4对。
②毛色 体背毛色一般为棕褐色,毛基深灰色,上段棕褐色,毛尖多为黑褐色,自头顶部至尾基部,沿背中央由黑色毛形成1条黑色纵纹。两颊部棕色,耳壳背面具棕黄色或棕褐色短毛,一般与体背同色。体腹面及四肢内侧为灰白色,毛基灰色,毛尖白色,体侧与体腹毛色界限分明。体侧较体背色淡。眼周为棕黄色。尾背腹两色,背方为暗棕色,腹方为淡灰色。尾毛稀疏而短,鳞片环清晰。四足背面被毛为白色,直达趾端。
③头骨 头骨略显细长,吻部较尖细,鼻骨狭长。额骨与顶骨间骨缝稍向后凸,但弧度甚小,故几近平直,使顶骨的外角近乎直角。顶间骨前缘呈“W”形,后缘略凸。听泡小而突出,门齿孔一般不达或仅及第1对上臼齿(M1)前缘连线(图4.54)。
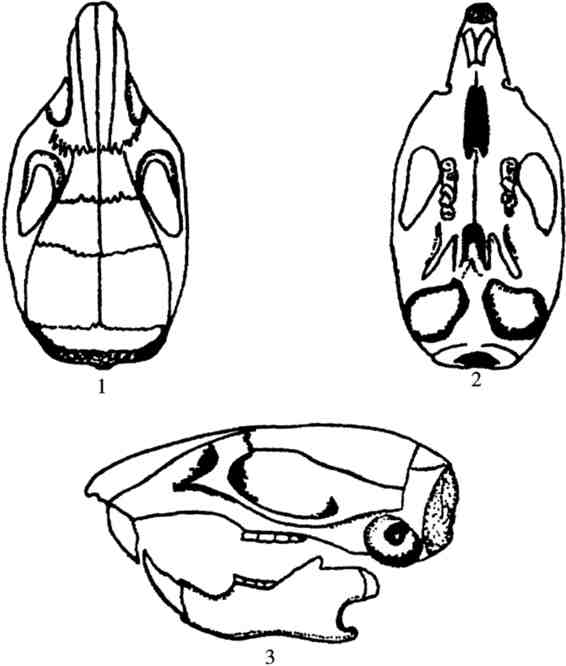
图4.54 黑线姬鼠的头骨
1.背面观;2.腹面观;3.侧面观
④牙齿 门齿唇面橙黄色。第1上臼齿(M1)咀嚼面具完整的3个横嵴,每一横嵴各具3齿尖,中央齿尖发达。第一横嵴外侧齿尖较小,内侧齿尖后移,呈新月形弯曲。第2上臼齿(M2)第一横嵴缺少外侧及中央齿尖,第二横嵴正常,第三横嵴的外侧齿尖小且与第二横嵴外齿尖相连。第3上臼齿(M3)小而近于圆形,咀嚼面分成两齿叶,内前方有一独立齿尖,可谓黑线姬鼠之特征。老年个体第3上臼齿(M3)咀嚼面不见上述两齿叶,而形成1个中部略凹的环形。
⑤量衡度 黑线姬鼠量衡度有关数据,见表4.29、表4.30。
表4.29 黑线姬鼠外形量衡度

表4.30 黑线姬鼠头骨量衡度 单位:mm

3)分类讨论 黑线姬鼠(Apodemusagrarius)自Pallas于1771年订名以来,其种级分类地位争议较少。
4)亚种分化 有关黑线姬鼠的亚种分化,尚有不同看法。夏武平(1984)认为黑线姬鼠在国内有5个亚种,即指名亚种(A.a.agrarius)、长江亚种(A.a.ningpoensis)、华北亚种(A.a.pallidior)、东北亚种(A.a.mantchuricus)和台湾亚种(A.a.insulaemus)。刘春生等(1991)提出中国大陆东部的黑线姬鼠可分为两个亚种,即上述的东北亚种和长江亚种。在秦岭-淮河以北地区者应归属东北亚种,而此线以南者应归属长江亚种。王廷正和许文贤(1993)则认为上述结论有待进一步研究,且华北亚种应暂予保留为妥,本书从此意见。
河南省分布有黑线姬鼠的2个亚种。
①华北亚种(A.a.pallidior,Thomas 1908)本亚种主要特征是体背中部黑色纵纹明显可见。分布于灵宝、济源、新乡、南阳等大部分地区。
②长江亚种(A.a.ningpoensis,Swinhoe 1870)本亚种主要特征是体背中央黑色纵纹不明显或隐约可见。分布于商城、固始、罗山等地。
5)地理分布 黑线姬鼠在河南的分布几乎遍布全省各地(图4.55)。国内除西藏、广东南部、海南以外,绝大多数省(区)均有分布。国外自朝鲜半岛经蒙古、俄罗斯直到西欧,都有分布。
6)生态 黑线姬鼠主要栖息于山麓地带的林缘、灌丛及草丛中,平原农作区较为潮湿的农田、菜地也大量分布。据1980年在兰考、孟津、新乡等地,1984年3月至1986年3月在遂平、中牟、周口等地,1994年4~10月在灵宝、新乡、镇平、商城、固始、罗山、济源等地调查,黑线姬鼠是河南省平原农作区的优势鼠种。在当地农田害鼠中所占比例平均为63%,高者可达90%以上。
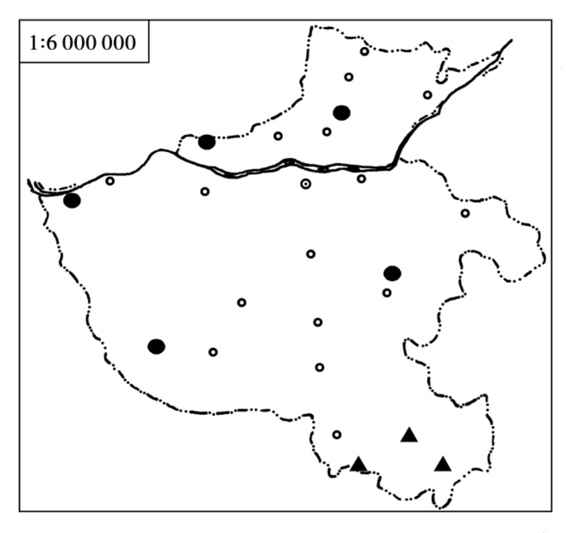
图4.55 黑线姬鼠的分布
●华北亚种(A.a.pallidior) ▲长江亚种(A.a.ningpoensis)
黑线姬鼠的洞穴结构比较简单,一般有洞口2~3个。巢室呈圆球形,以干草、作物茎叶、树叶等铺垫而成。洞道不深,长1~2 m,分2~4岔。黑线姬鼠食性较杂,但以植物性食物为主。食物常随季节不同而有变化,秋、冬两季以植物种子为主,辅以植物根茎;春天开犁播种后,则盗食种子和青苗;夏季取食植物的绿色部分及瓜果,并捕食昆虫。黑线姬鼠不冬眠,以夜间活动为主,黄昏与清晨最为活跃。春、秋两季活动频繁,常随自然条件变化和食物来源而迁移。
黑线姬鼠平均寿命1.5年左右。每年繁殖3胎左右,每胎产4~8仔,平均5.4仔,多者可达11仔。每年繁殖期为2~10月份。种群内雌雄性比的年间变化较小,平均为1∶1.08,雌性略少于雄性。黑线姬鼠种群数量的季节消长呈现双峰型。第一个高峰在5月份,捕获率可达9.9%,第二个高峰在10月份,捕获率为7.7%。全年怀孕率以4月份为最高,可达83.6%,次为9月份,达67.6%。
黑线姬鼠在春季的高捕获率,并非是繁殖的结果,而是由于气温升高,寻偶、觅食活动频繁,范围大。此时捕获者全是去年的成年个体和秋季即将进入繁殖的亚成体。只有在5月下旬才开始捕到当年出生的体重15 g左右的幼体。从6~12月份未成体比例增加,说明秋季的高捕获率是当年繁殖的结果。
7)经济意义 黑线姬鼠是豫东豫北平原、南阳盆地等广大农作区的重要害鼠之一。其危害特点是咬断作物茎秆,只取果实,取食数粒后,弃之又取他株。危害期自作物灌浆直至成熟。故对农业生产危害严重。同时,该鼠还是流行性出血热、地方性斑疹伤寒的带菌者和传染源。故应集中力量予以重点防控,以减少对农业生产的危害和降低对人民身体健康和生命安全的威胁。
(2)中华姬鼠(Apodemus draco,Barett-Hamilton 1900)
地方名:林姬鼠,龙姬鼠。
英文名:Chinese wood mouse
1)鉴别特征 上体毛色较大林姬鼠略暗,为暗黄褐色,耳壳较大,耳背为黑褐色。门齿孔较长,第1上臼齿(M1)后内尖发达,尾长略超过体长。
2)形态描述(图4.56)

图4.56 中华姬鼠
①外形 尾长略长于体长,尾鳞清晰。耳壳稍大,耳长平均为16 mm。耳前基部毛较长,形成毛丛。前足4指,有跖垫6枚,后足5趾,有跖垫6枚。后足长平均为20.7 mm,雌性具乳头3~4对。
②毛色 体背自头顶至尾基以及四肢外侧为灰褐色或黄褐色,毛基暗灰色,毛尖暗褐色。耳壳颜色较体背为深,呈棕黑或黑褐色。颊部和体侧毛色较体背略淡,但无明显界限。体腹面与四肢内侧为灰白色,毛基灰色,毛尖白色,与体侧毛色界限分明。尾背腹两色,背方暗褐色,腹方灰白色。足背呈污白色。
③头骨 头骨侧观背腹稍扁平,鼻吻部长而略尖。额骨与顶骨间的骨缝向后呈弧形凸出,额骨稍隆起。眶上嵴较明显。顶间骨略向下方倾斜。门齿孔略长,其后缘几乎达第1对上臼齿(M1)前缘连线(图4.57)。
④牙齿 第1上臼齿(M1)较大,具3个横嵴及3纵列齿尖,中列齿尖较大,两侧者较小,第三横嵴的舌侧齿尖较为发达。第2上臼齿(M2)第一横嵴中央齿尖消失,两侧者成为独立齿尖,第二、第三横嵴齿尖完全。第3上臼齿(M3)最小,内侧具3个齿尖。
⑤量衡度 中华姬鼠量衡度有关数据,见表4.31、表4.32。

图4.57 中华姬鼠的头骨
1.背面观;2.腹面观;3.侧面观
表4.31 中华姬鼠外形量衡度

表4.32 中华姬鼠头骨量衡度 单位:mm

3)分类讨论 Barrett-Hamilton于1900年把采自福建桂墩的一组标本订名为Mus sylvaticus draco。Allen(1912,1940)把其学名订为Modecus sylvaticus draco。Ellerman和Morrison-Scott(1951)把draco列为小林姬鼠(A.sylvaticus)的同物异名。小林姬鼠耳郭较小,头骨无眶上嵴,额骨与顶骨间的骨缝呈“V”形,分布于我国新疆及西藏西部。因此,Corbet(1978)把A.draco列为独立种,夏武平(1984)、王廷正和许文贤(1993)均认为A.draco应是一独立种,本书从此见解。
4)亚种分化中华姬鼠在国内的亚种分化,根据夏武平(1984)的意见应为3个亚种,即指名亚种A.d.draco、台湾亚种A.d.semotus、西南亚种A.d.orestes,而依冯祚建等(1986)记述,共有4个,即指名亚种(A.d.draco)、台湾亚种(A.d.semotus)、西南亚种(A.d.orestes)和川藏亚种(A.d.latronum)。其中,A.d.orestes与A.d.latronum形态差异较大,二者在四川、云南等省的广泛区域内有同域分布现象。因此,王廷正和许文贤(1993)提出,将川藏亚种(A.d.1atronum)订为大耳姬鼠(A.1atronum)较为妥当。
①指名亚种(A.d.draco,Barrett-Hamilton 1900)该亚种的模式产地为福建桂墩。其体背毛色较鲜明,尾长接近或略长于体长;耳较短,为14~17 mm;后足较短,20~23 mm;后足踝部通常暗色。本亚种分布于中国东部南方各省,在北方分布于陕西、甘肃、宁夏、河北、北京等地。
②西南亚种(A.d.orestes,Thomas1911)该亚种的模式产地为四川峨眉山。其体背毛较暗;尾长长于体长;耳较大,为17~18.5 mm;后足较长,23~25 mm;后足踝部白色。该亚种分布于四川、贵州、云南、西藏东部及陕西等地。在国外见于缅甸、印度东北部。
③台湾亚种(A.d.semotus,Thomas 1908)该亚种的模式产地为台湾阿里山。其体背、腹部及四足毛色均很深,该亚种分布于台湾。
河南省分布有1个亚种——指名亚种(Apodemus draco draco),本亚种主要特征是尾长接近体长,其余特征见种级记述。见于灵宝等地。
5)地理分布中华姬鼠在河南省分布于豫西黄土台地、豫西伏牛山地等(图4.58)。国内分布于福建、台湾、四川、云南、西藏、陕西、甘肃、黑龙江、河北、山西、湖北、宁夏等省(区)。国外分布于缅甸及印度东北部。
6)生态中华姬鼠栖息于亚热带、暖温带和寒温带阔叶林、针阔叶混交林及灌丛中,属喜湿性鼠类。常在林间石头下、倒木旁、树根中、苔藓层中居住。多在夜间活动,白天少有出没。以植物的种子和绿色部分为主食。繁殖期为6~9月份,每胎产2~4仔。据1994年4月在灵宝调查,捕获率为1.6%。
7)经济意义 中华姬鼠在农作区危害农作物种子和幼苗,在林区可危害苗木,但目前在河南省数量较少,分布范围亦较狭窄。

图4.58 中华姬鼠的分布(▲)
(3)大林姬鼠(Apodemus peninsulae,Thomas1906)
地方名:林姬鼠。
英文名:Large field mouse
1)鉴别特征 体型较中华姬鼠稍大,体背黄褐色或棕黄色。足背面灰色。尾背腹两色,尾长多短于体长。门齿孔较短,其末端距第1上臼齿(M1)前缘连线较远。
2)形态描述(图4.59)

图4.59 大林姬鼠
①外形 体型中小,尾长一般短于体长。尾毛不发达,尾上鳞环清晰。耳壳小而薄,耳长一般不超过18 mm。前足掌部裸露,具掌垫6枚;后足跖部也裸露,具跖垫6枚。雌性具乳头4对。
②毛色 体背毛色较中华姬鼠为淡,体背从头至尾基以及四肢外侧均为黄棕褐色,毛基灰色,上段棕黄色,毛尖黑色,并杂生一些全黑色毛。耳壳与体背颜色基本一致。两颊和体侧为黄褐色,与体背几乎无明显界限。颏部纯白色。喉、体腹面及四肢内侧灰白色,毛基灰色,上段白色。足背白色,尾背腹两色,背方棕褐色,腹方白色。冬毛较夏毛颜色浅淡。
③头骨 头骨椭圆形,吻部略显圆钝,鼻骨前端膨大。额骨与顶骨间骨缝向后呈圆弧形凸出,额骨之眶间部稍隆起,眶上嵴明显,顶间骨宽短且略向后倾斜。颧弓不明显向外扩张。枕骨比较陡直。听泡较小。门齿孔后缘不达上臼齿列前缘连线(图4.60)。

图4.60 大林姬鼠的头骨
1.背面观;2.腹面观;3.侧面观
④牙齿 门齿唇面橙黄色。第1上臼齿(M1)特大,约为第2、第3上臼齿(M2、M3)之和。第1上臼齿(M1)咀嚼面上有3个横嵴。每一横嵴均由3个齿尖构成,中央齿尖均很发达,形成3个不同程度的新月形。第三横嵴的舌侧齿尖退化或较小。第2上臼齿(M2)亦有3个横嵴,第一横嵴无中央齿尖,舌侧齿尖较大,唇侧齿尖较小。第3上臼齿(M3)最小,内侧具3个齿尖。
⑤量衡度大林姬鼠量衡度有关数据,见表4.33、表4.34。
表4.33 大林姬鼠外形量衡度

表4.34 大林姬鼠头骨量衡度 单位:mm
3)分类讨论 Thomas于1906年将朝鲜半岛的此类姬鼠订名为Micromys specious peninsulae(日本姬鼠朝鲜亚种)。Allen(1940)则认为日本姬鼠的个体较大,其分布地仅限于日本列岛,故将亚洲大陆的大林姬鼠确定为一个独立的种,订正为Apodemus peninsulae。此后,仍有许多学者将其作为日本林姬鼠(A.specious)或黄喉姬鼠(A.flavicoliis)的一个亚种。据资料记述,A.speciou的个体大,头骨较长,最大头骨长达33.5 mm。后足较为发达。与A.peninsulae在毛色上也有差异。Jones(1956)把丹麦、瑞典等欧洲国家的黄喉姬鼠(A.flavicollis)标本与朝鲜的大林姬鼠(A.peninsulae)标本进行了比较,发现前者具有黄色喉斑,雌性多具乳头3对;门齿孔较长,与上臼齿列之前缘几乎在同一水平线上;M3较小,诸特征均有别于后者,故再次提出A.peninsulae应是独立种。另外,Allen(1940)、Zimmermann(1964)、Corbet(1978)、夏武平(1984)、郑昌琳(1986)、王廷正和许文贤(1993)等学者均认为A.peninsulae应是独立的种。尽管一些俄罗斯学者坚持认为分布于东亚大陆和日本列岛上的均为日本姬鼠(A.speciosus),但总体来看,Tokuda(1941)将其作为Apodemus ainu的亚种及Ellerman&Morrison-Scott(1951)将其列为Apodemus flavicollis的亚种,都是欠妥当的。
夏武平(1984)对中国的姬鼠属动物的分类体系重新进行修订,并进一步探讨了其与日本所产姬鼠的关系。夏武平指出,应将大林姬鼠(A.peninsulae)作为独立的种;日本姬鼠(A.speciosus)的M2之前外齿突较大林姬鼠更为退化,有可能是从后者发生变化而形成的物种。大林姬鼠分布到朝鲜及西伯利亚,在西伯利亚通过库页岛进入日本的北海道,在朝鲜的则可能经马岛到达日本,后发展成日本姬鼠。但是,中国仍有不少学者沿用A.speciosus给中国的大林姬鼠命名。
4)亚种分化大林姬鼠已记述的亚种计有5个,国内有3个亚种即华北亚种(A.p.sowerbyi,Jones1956)、东北亚种(A.p.praetor,Miller 1914)和青海亚种(A.p.qinghaiensi,Feng,Zheng etWu 1983)。
①东北亚种(A.p.praetor,Miller 1914)该亚种的模式产地为吉林省吉林市西南部。其体型较大;听泡较长,平均为5.5 mm。该亚种主要分布于东北三省及内蒙古东部。
②华北亚种(A.p.sowerby,Jones 1956)该亚种的模式产地为山西北部桂华城。其体型较东北亚种略小,听泡平均长为5.0 mm。该亚种分布于华北及西北东部,包括山东、河北、天津、北京、陕西、山西、甘肃、宁夏和内蒙古等省(区)。
③青海亚种(A.p.qinghaiensis,Feng,Zheng etWu 1983)该亚种的模式产地为青海乐都。其体型较东北亚种小,听泡小且低平,平均长度为4.5 mm,该亚种分布青海东部、四川西部和西藏东部等地。
河南省分布有1个亚种——华北亚种(A.p.sowerbyi,Jones 1956)本亚种主要特征是听泡大而鼓胀,甚为发达,平均长度为5 mm。体型较小。见于济源等地。
5)地理分布 大林姬鼠在河南省分布于豫西北太行山地、豫西伏牛山地等(图4.61)。国内分布于东北、西北、华北及西南诸省(区)。国外分布于自朝鲜至西伯利亚的国家和地区。
6)生态 大林姬鼠主要栖息于山区、高原地带的林区及草甸灌丛中。喜栖于湿润型或半湿润型环境,是典型的森林鼠类。常在树根、倒木、灌丛或草丛等处掘土打洞。昼夜均活动,但以夜间活动为主。喜食植物种子、果实,也食少量昆虫及植物的绿色部分。每年繁殖1~4胎,每胎产2~11仔,平均5.7仔。繁殖期为4~8月份,以5~6月份最盛。由于繁殖的季节性强,故其种群数量的季节性消长极为明显,春季最低,秋季最高。
7)经济意义 大林姬鼠在林区数量较多,喜食针、阔叶树的种子和果实,亦取食嫩叶、幼苗。故对森林的自然更新及人工造林危害较大,应注意加强防治措施。另一方面,大林姬鼠储食行为与林木更新的关系尚待进一步探讨。

图4.61 大林姬鼠的分布(▲)
4.3.4.2小鼠属Mus
小家鼠(Musmusculus,Linnaeus1758)
地方名:耗子、小老鼠。
英文名:Housemouse
(1)鉴别特征 体小型,体长不及100 mm。体毛柔软,尾长短于或近于体长。上颌门齿末端内侧具明显缺刻。体背毛色为黄褐色或灰褐色。
(2)形态描述(图4.62)

图4.62 小家鼠
1)外形 小型鼠类,体长70 mm左右,尾长略短于体长,鳞环可见。吻部较长而尖。四肢细弱,前足掌垫5枚,后足跖垫6枚。雌性具乳头5对。
2)毛色 头部、体背部呈灰褐色或棕褐色,毛基部灰色,毛尖褐色或棕色。杂生较多的黑色长毛。体腹面毛白色,多数个体毛基、毛尖均为白色,有些毛基灰色或淡灰色,毛尖白色。体侧与体腹毛色界限分明,耳壳深褐色,内外面均被以褐色短毛。尾背腹两色,背方淡褐色,腹方污白色。前后足足背纯白色或污白色。
3)头骨 颅骨呈长椭圆形,且较低平。鼻吻部较短而厚,侧观头骨背面稍隆起,表面光滑,眶上嵴和颞嵴不发达,眶间宽小于4 mm。顶间骨宽短,宽约为长的3倍。颧弓较细弱,不明显向外扩张,门齿孔狭长,其后缘超过第1上臼齿(M1)前缘连线,听泡小而略扁平(图4.63)。

图4.63 小家鼠的头骨
1.背面观;2.腹面观;3.侧面观
4)牙齿 上门齿唇面橙黄色,齿冠内侧具一明显缺刻,下门齿齿尖恰嵌于此。第1上臼齿(M1)发达,约相当于或略大于第2、第3上臼齿(M2、M3)之和。咀嚼面具3个横嵴,第一、第二横嵴舌侧齿尖位置偏后,并向内弯曲成月牙状,第三横嵴缺内侧齿尖。第2上臼齿(M2)的第一横嵴仅留内侧齿尖,余者皆退化。第二横嵴的3个齿尖发育正常,第三横嵴缺内侧齿尖。第3上臼齿(M3)很小,具3个齿尖。第1下臼齿(M1)咀嚼面具三个横嵴,均由2个齿尖构成;第2下臼齿(M2)具二横嵴,各有2个明显齿尖;第3下臼齿(M3)也有二横嵴,第一横嵴有2个齿尖,第二横嵴具1个齿尖。
5)量衡度 小家鼠量衡度有关数据见表4.35、表4.36。
表4.35 小家鼠外形量衡度

表4.36 小家鼠头骨量衡度 单位:mm

(3)分类讨论 小家鼠(Musmusculus)自Linnaeus于1758年订名以来,种级分类地位比较稳定,没有多少争议。
(4)亚种分化小家鼠在全世界广泛分布,故有关其亚种分化十分复杂,学者们意见颇不一致,已记述的亚种达60个。各家所描述的亚种多以变异较大的毛色等外部特征为鉴别依据,故已订名的亚种同物异名者甚多。根据Ellerma和Morrison-Scott(1951)的意见,中国的小家鼠分化为5个亚种,即华东亚种(M.m.castaneus)、喜马拉雅亚种(M.m.homouru)(原川陕亚种M.m.tantillus为其异名)、西南亚种(M.m.urbanus)、西北亚种(M.m.wagneri)(原甘肃亚种M.m.gansuensis、北疆亚种M.m.decolor和南疆亚种M.m.pachycercus均为其异名)和东北亚种(M.m.manchu)。Johes和Johnson(1965)则提出了不同意见。Marshall(1977,1981,1986)、Marshall和Sage(1981)先后对世界各地的小家鼠进行了系统的比较研究,综合考虑形态、生态、染色体组型、同工酶等方面的研究结果,对小家鼠的同物异名、亚种划分等进行细致整理,其最终结果与Ellerma和Morrison-Scott(1951)的结果非常相近。据Marshall(1986)的意见,中国的小家鼠共有4个亚种,其与Ellerma和Morrison-Scott(1951)的不同之处在于,Marshall认为小家鼠西南亚种(M.m.urbanus)=小家鼠华东亚种(M.m.castaneus);将小家鼠东北亚种(M.m.manchu)和韩国亚种(M.m.yamashinai)均归入日本亚种M.m.molossinus中;并指出采自四川的小家鼠川陕亚种(M.m.tantillus)标本实际上为日本亚种molossinus,为后者与华东亚种的过渡类型。目前比较公认的是小家鼠在国内约有7个亚种。
1)华东亚种(M.m.castaneus,Waterhouse 1834)该亚种模式产地为菲律宾群岛。其尾较长,多数仅略短于体长;体色较深,背毛呈棕灰色或棕褐色,基部深灰色,毛尖棕色;腹毛多呈土黄色或暗褐色,毛基深灰色;背毛、腹毛在体侧分界不明显;尾二色不显著,尾上棕褐色,尾下为土黄色;四足与背同色,仅趾尖为白色;颧板宽,其前缘向前倾斜,此为该亚种的显著特征。该亚种以家栖为主。在中国分布于华东沿海一带以及湖北、云南、贵州、福建和台湾等省,湖南、广西、广东、江西可能也有分布。国外见于南亚和东南亚的大部分地区及日本、东非、南非等地。
2)喜马拉雅亚种(M.m.homourus,Hodhso 1845)该亚种的模式产地为尼泊尔。其体型相对较大,平均体重约16 g。尾长几与体长相等。体色较暗,背部暗褐色,体侧赭黄色较著;腹部呈灰白色,毛基浅鼠灰色,毛尖白色;尾二色显著;足背色浅,白色或黄白色。颧板前缘直,不向前凸突成半圆形。该亚种主要为户外型。在中国分布于西藏、云南、广西、四川、陕西、甘肃、湖北等地。国外见于尼泊尔、锡金、克什米尔、巴基斯坦等地。
3)北方亚种(M.m.wagneri,Eversmann 1848)该亚种的模式产地为俄罗斯伏尔加河与乌拉尔河之间的Kamysh-Samarian湖畔。尾长明显短于体长,约为体长的70%。体色较浅,背部淡褐色或黄褐色或沙土色,腹部纯白色或灰白色,背毛、腹毛在体侧界限分明;尾二色但不甚显著,尾上淡褐色,尾下污白色;四足背面近白色。颧板前缘向凸突成弧形或半圆形。该亚种为户外型。在中国分布于新疆、内蒙古、青海、西藏、甘肃、陕西、山西、河北、北京、山东等地。国外见于中亚和蒙古。
4)日本亚种(M.m.molossinus,Temminck 1845)该亚种的模式产地为日本。其尾长明显短于体长。体背部为深灰褐色至黑褐色,背中央分布有很多全黑色毛;腹部白色或灰白色,毛基灰白色,背毛、腹毛在体侧的分界不甚明显;尾二色,尾上为棕褶色,尾下近白色;四足背面白色或近白色。颧板前缘明显凸突呈半圆形。该亚种主要为户外型。在中国分布于黑龙江、吉林、辽宁、内蒙古东南部、河北东北部、山西和四川的部分地区。国外见于朝鲜、韩国、日本和西伯利亚等。
此外,Corbet和Hill(1992)、黄文几等(1995)认为中国尚有指名亚种(M.m.musculus)的分布。该亚种的尾长短于体长,体背面为灰黄褐色,腹部为棕灰色;尾二色,后足褐色。颧板前缘直。该亚种主要为室内型。在中国分布于沿海大中城市,在许多地方还与本地亚种相混杂,故毛色变化较大。国外分布于欧洲北部和东部、西伯利亚西部、朝鲜等地。
河南省分布有2个亚种。
北方亚种(M.m.wagneri),本亚种主要特征是尾长短于体长,体背毛色为一致的沙黄色,体腹毛色由毛基至毛尖均为纯白色。分布于豫西灵宝、新乡、镇平、济源等地。
华东亚种(M.m.castaneus),本亚种主要特征是尾长接近于体长,体腹面毛色呈暗褐色或土黄色,尾背腹两色不明显。分布于商城、罗山、新县等地。
(5)地理分布小家鼠在河南省除海拔较高地区之外,各地均有分布,属广布种(图4.64)。国内分布几遍全国各地。从世界范围来看,除了南极和一些海洋岛屿、亚寒带针叶林和苔原带边缘外,几乎全球都有其踪迹。

图4.64 小家鼠的分布
●北方亚种M.m.wagneri ▲华东亚种M.m.castaneus
(6)生态小家鼠为与人类伴生的家野两栖小型啮齿动物,适应能力强,栖息环境非常广泛。在建筑物中,从仓库、厨房、居室、办公室、楼房,以至于禽舍畜圈中都有其分布。在野外,小家鼠主要栖息于农田、菜地、荒地、林缘、灌丛、草丛等处。小家鼠营巢的巢材很广,包括纸屑、破布、棉絮、毛、羽、杂草、植物叶子等。野外常有两种洞穴,一种为临时洞,多为盲洞,洞道较短,多为躲避天敌及临时休息之用。另一种为居住洞,有洞口2个以上,洞口直径2~3 cm。有时尚有一个通气洞,由地面直达巢室。雌鼠产仔后,将大部分洞口堵住,只留1个洞口。小家鼠昼夜活动,但以夜间活动为主。全天有两个活动高峰,即黄昏后和黎明前。小家鼠还具有明显的季节性迁移现象,春季由室内迁往野外,晚秋季随作物收获完毕,灌丛、草丛逐渐枯萎,又大量迁回居民区建筑物和秸秆垛中,并于此越冬。小家鼠食性较杂,以盗食粮食作物和经济作物为主。如小麦、玉米、水稻、花生等。春季食源贫乏时,也咬食麦苗。夏季在野外也食草籽和昆虫,数量大时还啃食树皮、棉桃、瓜果等。其食性随季节、栖息环境、食源等的不同而略有变化。
小家鼠的繁殖能力极强,如果条件适宜,几乎全年均可繁殖。以夏、秋两季为繁殖盛期,全年一般产6~8胎,多达10余胎,孕期平均18~20 d,每胎产6~9仔,多者可达10余仔。幼鼠出生2个月即达性成熟。
小家鼠的自然寿命平均为1年。据1984年3月至1986年3月在遂平、中牟、周口等地调查,在家栖鼠类中,小家鼠可占14.4%~20.4%,平均为15.6%。在野外则占0.62%~6.3%,平均为3.02%。家栖者其种群数量的季节消长呈现双峰型,第一个高峰在7月份,第二个高峰不明显,较第一高峰为低并延至12月份。形成第一个高峰,一方面是由于春季繁殖的结果,另一方面是因为天气炎热,小家鼠在野外活动减少而较多地在居民区活动。形成第二个高峰主要是由于农作物已经成熟收获完毕,气候逐渐变冷,野外食源趋于贫乏,小家鼠逐渐迁入室内。
(7)经济意义 小家鼠盗食粮食,而且数量大,分布极广,故为重要农业害鼠之一。由于伴人栖居,常常咬坏衣物、家具、书籍,并盗食、污染食品。此外,还携带和传播流行性出血热等自然疫源性疫病,威胁人类身体健康。同时,对城乡居民建筑物、电力设备,通信设施等也时有破坏,故各行各业应齐心协力,加强对小家鼠的灭防工作。
4.3.4.3 家鼠属Rattus
河南省家鼠属分种检索表

(1)褐家鼠(Rattus norvegicus,Berkenhout1769)
地方名:大家鼠。
英文名:Common Rat or Bwown Rat
1)鉴别特征 体型粗壮,尾短粗,尾长明显短于体长,但超过体长的2/3。耳短圆而厚。头骨两颞嵴近于平行。
2)形态描述(图4.65)
①外形 体型粗壮,尾短粗,尾长短于体长,尾毛短而稀疏,鳞环外露明显。耳短圆而厚,前折不达眼部。后足粗壮、坚实,其长度为27~38 mm,前足垫5枚,后足垫6枚。雌性有乳头6对,胸部2对,腹部1对,鼠鼷部3对。
②毛色 褐家鼠毛色随年龄、栖息环境的不同而有所变化,但季节性变化不大。通体毛色棕褐色至灰褐色或暗褐色,毛基灰色,毛尖棕褐色。头及体背尤以体背中部杂生较多的黑色长毛,使背中央颜色较深。体侧的黑色长毛较少。体腹面灰白色,毛基灰色,毛尖污白色,体侧与体腹面毛色界限分明,尾背腹两色,背方黑褐色,腹方灰白色。四足足背白色。幼体毛色较深,体背黑灰色,毛尖棕色极少,而且细软。③头骨头骨整体观粗壮而略长,吻部较大,鼻骨略长而宽。眶上嵴发达,并与发达的颞嵴相连,向后延伸至鳞骨,两侧颞嵴近平行。颧弓坚实。顶间骨宽度约等于二顶骨宽度之和。听泡中等大小。门齿孔后缘接近上臼齿列前缘连线(图4.66)。

图4.65 褐家鼠
④牙齿 第1上臼齿(M1)发达,咀嚼面具3行横嵴,第一横嵴外侧齿尖略有痕迹,第二横嵴的齿尖极退化,外侧齿尖略明显,第三横嵴内侧齿尖消失。第2、第3上臼齿(M2、M3)的第一横嵴均退化,仅存一内侧齿尖。第2上臼齿(M2)的第二、第三横嵴上的齿尖尚清晰。第3上臼齿(M3)的第二、第三横嵴已融合,呈马蹄形齿环。第3上臼齿(M3)最小,约为第1上臼齿(M1)之半。下颌臼齿咀嚼面的齿尖不明显,但横嵴尚较清楚。
⑤量衡度 褐家鼠量衡度有关数据,见表4.37、表4.38。
3)分类讨论褐家鼠(Rattusnorvegicus)自Berkenhout于1769年订名以来,其种级分类地位比较稳定,争议主要集中于亚种分化上。
4)亚种分化 吴德林(1982)研究了中国23个省(区)的标本,认为除台湾省外,中国的褐家鼠已分化出4个亚种,即指名亚种(R.n.norvegicus Berkenhout)、甘肃亚种(R.n.soccer Miller)、华北亚种(R.n.humiliates Milne-Edwards)和东北亚种(R.n.caraco Pallas)。罗蓉(1993)指出,贵阳的褐家鼠标本中一些测量数值超过吴德林(1982)报道的甘肃亚种的最高值、也超过指名亚种的最低值,因此,有关贵州的褐家鼠的亚种分化问题尚需进一步探讨。
①指名亚种(R.n.norvegicus,Berkenhout1769)该亚亚种的模式产地为英国。其体型较大,体长162~260 mm,平均220 mm;尾长135~230 mm,平均193 mm;后足长38~45 mm,平均41 mm。被毛较粗糙。该亚种分布于东南沿海及附近岛屿,包括广东、海南、厦门等地,系过去由外轮传入。
②华北亚种(R.n.humiliates,Milne-Edwards 1868)该亚种的模式产地为北京附近。其体型在4个亚种中最小,体长122~215 mm,平均165 mm;尾长84~165 mm,平均130 mm;后足长25~66mm,平均30mm。该亚种分布于淮河以北、太行山以东地区,北至内蒙古高原、东北方向可达辽东半岛。
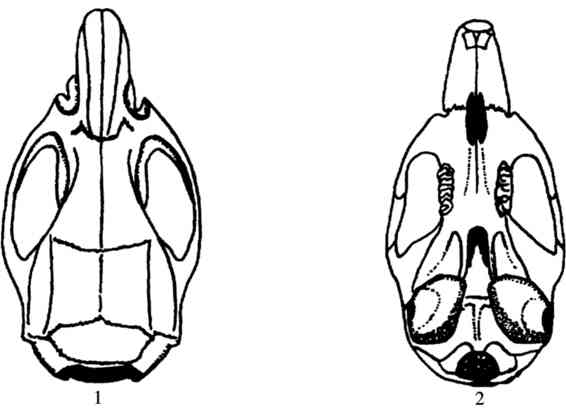

图4.66 褐家鼠的头骨
1.背面观;2.腹面观;3.侧面观
表4.37 褐家鼠外形量衡度
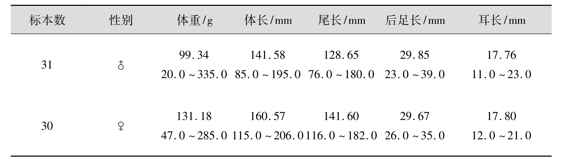
表4.38 褐家鼠头骨量衡度 单位:mm
 (https://www.xing528.com)
(https://www.xing528.com)
③甘肃亚种(R.n.socer,Miller 1914)该亚种的模式产地为甘肃(Taochow)。其体型中等,体长142~258 mm,平均187 mm;尾长105~211 mm,平均153 mm;后足长31~34 mm,平均38 mm。该亚种分布于淮河流域以南、太行山以西地区,西至甘肃、青海、四川,南抵云南和广西,北达内蒙古地区。
④东北亚种(R.n.caraco,Pallas 1779)该亚种的模式产地为西伯利亚东部外贝加尔地区(Transbikalia)。体型中等,体长135~250 mm,平均188 mm;尾长105~180 mm,平均147 mm;后足长28~39 mm,平均34 mm。该亚种分布于黑龙江、吉林、辽宁、内蒙古;国外见于俄罗斯、朝鲜和日本。
河南省分布有两个亚种。
华北亚种(Rattus norvegicus humiliates),本亚种的主要特征是成体体长平均在165 mm左右,后足长平均30 mm左右,见于新乡、镇平等地。
甘肃亚种(Rattus norvegicus soccer),本亚种的主要特征是体型中等,成体体长平均在185 mm以上,后足长平均在34 mm以上,见于灵宝等地。
5)地理分布 褐家鼠在河南省分布遍及全省各地(图4.67),国内除西藏以外,广布于全国各地。褐家鼠很可能起源于黑龙江流域和西伯利亚东南部。由于其繁殖力强,适应性极强,分布区逐渐扩大;同时与人类关系密切,通过火车、轮船甚至飞机等现代交通工具而不断扩散。迄今,除极地及严酷的荒漠地带外,几乎遍及全球,成为世界上分布最为广泛的啮齿动物。
6)生态 褐家鼠可谓世界上分布最广泛的啮齿动物,常伴人而居,栖息于城乡住宅、仓库、厨房、厕所、畜圈、下水道、农田、荒地、林缘、沟渠、河湖沿岸等多种生态环境,甚至火车、轮船、飞机等处都可见其踪迹。褐家鼠善于攀缘、游泳、掘土,洞道结构比较复杂。洞口多在墙角下或阴沟中,一般有洞口2~4个,但进口通常只有1个。出口处常有颗粒状松土,洞道长50~210 cm,分支较多。洞道距离地面深达150 cm,洞内有巢室1个,巢材为破布、破棉絮、碎纸、植物叶子等。褐家鼠食性很杂,几乎无所不食,但嗜食肉类及含水分较多的果品。在住宅区,主要盗食粮食和各种食品,在食源贫乏时,也吃垃圾、粪便;在野外条件下,主要以各种成熟的作物种子如水稻、花生、甘薯等为食,也取食植物的绿色部分、草籽、小鱼、虾、青蛙,甚至小鸡、小鸭等家禽。褐家鼠昼夜活动,但以夜间活动为主,清晨和黄昏是其活动高潮。视觉较差,但嗅觉、听觉、触觉均很灵敏。记忆力强,警惕性高,多沿墙根壁角行走,小心谨慎,对环境的改变十分敏感,性情凶暴,遇到紧急情况,来不及躲避时,即立起前身,发出叫声,扑向来者。
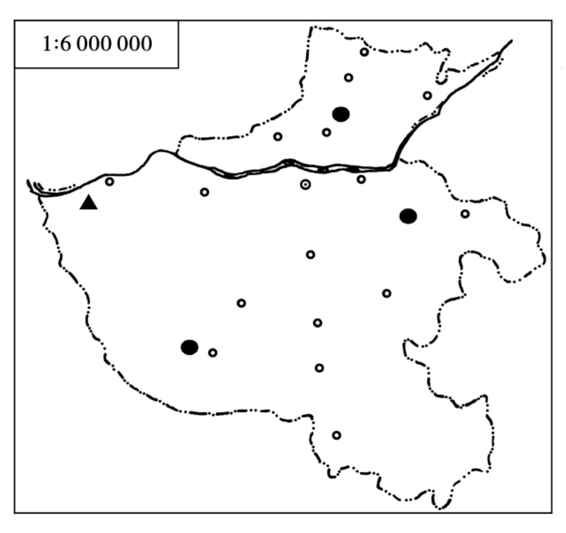
图4.67 褐家鼠的分布
●华北亚种R.n.humiliates ▲甘肃亚种R.n.soccer
褐家鼠繁殖能力很强,条件适宜时,全年皆可繁殖。每年可繁殖6~10胎,妊娠期约21 d,每胎产5~14仔,平均为7.4仔。褐家鼠平均寿命2~3年。据调查(王学高等,1980;葛荫榕等,1987),褐家鼠在豫东豫北平原室内鼠类组成中占67.8%~79.6%,平均为72.9%,占绝对优势。1994年4月在灵宝调查时,使用5只鼠夹,一夜间捕获9只成年个体。褐家鼠种群数量的季节消长呈现明显的双峰型。第一个高峰在7月份,第二个高峰在11月份。全年怀孕率最高在4月份,达81.8%,最低在2月份,为16.7%。全年平均怀孕率为47.7%,两次数量高峰恰好在二次繁殖高峰之后的两个月出现。捕获率上升月份与当年生未成体比例增多的月份相吻合,说明褐家鼠进入繁殖盛期早,持续时间长。因为栖息地和食源相对稳定,两个捕获率高峰都比较真实地反映了繁殖增长的结果。褐家鼠种群的周年雌雄性比为1∶0.79,雌性略多于雄性。
7)经济意义褐家鼠为农业、工业、卫生重要害鼠之一,盗食粮食,污染食品,危害农作物,破坏电力设施、通信设备,咬坏多种物资,又常咬伤幼畜雏禽,偶尔还咬伤婴幼儿。同时,尚可携带和传播鼠疫和流行性出血热等自然疫源性疾病。总体来看,褐家鼠对工农业生产、食品工业发展以及人类健康事业都有极为严重的危害和威胁。故在城乡各地均应将其列为重点对象,进行大力防治。褐家鼠毛皮尚有一定利用价值。
(2)大足鼠(Rattus nitidus,Hodgson 1845)
地方名:家鼠、灰胸鼠。
英文名:Himmalayan rat
1)鉴别特征 体型与褐家鼠相似,但后足较长而宽大,尾较长,几接近体长,耳壳大而薄,前折可达眼部。
2)形态 描述(图4.68)

图4.68 大足鼠
①外形 体型中等大小,近似于褐家鼠,但尾长接近体长,尾上鳞环不如褐家鼠的明显。后足较宽大而长,平均长度在34 mm左右。耳壳长而薄,前折可达眼部。雌性具乳头4对。
②毛色 体背部为暗棕褐色,略带黄色色调,毛基灰色,毛尖棕黄色,杂生棕褐色和黑色长毛,黑色长毛多集中于体背中央。体侧毛色稍淡。体腹面为灰白色,毛基灰色,毛尖白色或渲染微黄色。耳壳背面毛色深暗,尾单色,背方稍暗。前足足背白色,指亦白色。
③头骨 头骨侧观背腹略扁平而狭长,鼻骨窄长,向前可超出前颌骨前缘1 mm左右,向后延伸至前颌骨与额骨骨缝之后。眶上嵴发达,向后与颞嵴相连,并延伸至顶骨外角处。两颞嵴在头骨背面呈弧形,顶间骨近椭圆形,与上枕骨中部相连处几成垂直状,听泡突出。门齿孔末端略超过上臼齿列前缘的连线(图4.69)。
④牙齿 第1上臼齿(M1)具3列横嵴,第一横嵴唇侧的齿突较为退化,第二横嵴的3个齿尖正常,第三横嵴缺舌侧齿尖。第1上臼齿有7个齿尖,第2上臼齿(M2)的第一横嵴仅存舌侧齿尖,第二横嵴发育正常,而第三横嵴缺舌侧齿尖。第3上臼齿(M3)与第2上臼齿相似,但形小且三齿叶。下颌臼齿的齿尖均为二纵列。
⑤量衡度 大足鼠量衡度有关数据,见表4.39、表4.40。

图4.69 大足鼠的头骨
1.背面观;2.腹面观;3.侧面观
表4.39 大足鼠外形量衡度


表4.40 大足鼠头骨量衡度 单位:mm

3)分类讨论 大足鼠(Rattus nitidus)自Hodgson(1845)订名以来,其种级分类地位比较稳定。该种在外形酷似黑家鼠(R.rattus)的暗腹类型和拟家鼠(R.rattoides)。其区别于后两种鼠的主要特征在于鼻骨较狭长,向后延伸到前颌骨与额骨骨缝处或超过之。
4)亚种分化根据Ellerman和Morrison-Scott(1951)的意见,大足鼠已记述有3个亚种,即指名亚种(R.n.nitidus)、华北亚种(R.n.humiliatus Milne-Edwards)和泰国亚种(R.n.rahensis Kloss)(1918)。国内仅有指名亚种,但王廷正和许文贤(1993)认为,大足鼠在国内的亚种分化尚需进一步整理。
河南省目前分布有1个亚种。
指名亚种(R.n.nitidus,Hodgson 1845)本亚种主要特征见种级记述。分布于商城、罗山等地。
5)地理分布 大足鼠在河南省分布于豫南大别山地(图4.70)。国内分布于华东、华南、西南及陕西、西藏等地。国外分布于泰国、缅甸、尼泊尔等国家。

图4.70 大足鼠的分布(▲)
6)生态 大足鼠主要栖息于低山丘陵地带,营巢于田埂、溪流、水沟旁及草垛下。常在水稻、玉米、豆类田中觅食。食物以植物种子为主,喜食稻谷、玉米,也食浆果、草籽及小鱼、虾等。以夜间活动为主,白昼也见有出没。每年4~5月份为繁殖盛期,平均每胎产4~6仔。
7)经济 意义大足鼠为豫南大别山区主要农业害鼠之一,给当地农业生产造成一定危害,故应予以灭杀。有关其寄生虫学和流行病学意义需要深入调查研究。
(3)黄胸鼠(Rattus flavipectus,Milne-Edwards 1871)
地方名:黄腹鼠、长尾巴老鼠。
英文名:Buff-breasted rat
1)鉴别特征 体型中等,体形修长。尾长超过体长。耳大而薄,前折可遮住眼。体腹面毛呈浅黄色。前足背有一黑色纵纹。
2)形态描述(图4.71)

图4.71 黄胸鼠
①外形 体型中等,外观较褐家鼠修长,尾长明显大于体长,尾上鳞环可见。耳壳大而薄,前折可遮住眼部。后足较为细长,长度在30~37 mm之间。雌性有乳头5对,胸部2对,鼠鼷部3对。少数个体在腹部尚有1对。
②毛色 体背面毛为棕褐色,毛基深灰色,毛尖黄褐色或棕褐色。体背中央杂生较多的黑褐色长毛,故背中部颜色较体侧略深。体腹毛色灰黄,胸部黄色较浓,毛基灰色,毛尖浅黄色。前足背面中央呈暗灰褐色,周围灰白色;后足背面污白色。喉、四肢下部及肛门周围毛色较浅。尾上被有淡灰褐色细毛,鳞环清晰。幼体毛色灰黑,被毛质地柔软。
③头骨 不若褐家鼠者粗壮,吻部较短,鼻骨较长。颧弓略显细弱。眶上嵴发达并与颞嵴相连,后延至鳞骨。左右颞嵴均呈弧形向外弯曲,听泡较发达。门齿孔较长,其后缘可达上臼齿列前缘连线(图4.72)。
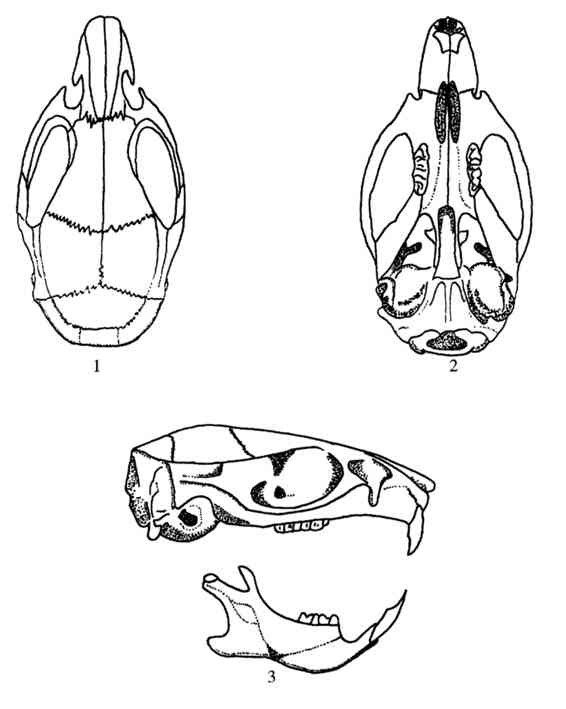
图4.72 黄胸鼠的头骨
1.背面观;2.腹面观;3.侧面观
④牙齿 第1上臼齿(M1)齿冠前有一带状隆起,咀嚼面具3行横嵴。第一、第二横嵴各具3个齿尖,内侧的2个齿尖位置略靠后。第三横嵴内侧齿尖退化,其余者正常。第2上臼齿(M2)第一横嵴仅有内侧齿尖,第二横嵴正常,第三横嵴形成1个齿尖。第3上臼齿(M3)较小,第一横嵴仅余1个内侧齿尖,其余则愈合成1个“C”形齿环。下颌3个臼齿咀嚼面齿尖不甚明显,但横嵴尚清晰。第1、第2、第3下臼齿(M1、M2、M3)咀嚼面分别具有3、2、2个横嵴。
⑤量衡度 黄胸鼠量衡度有关数据见表4.41、表4.42。
表4.41 黄胸鼠外形量衡度

表4.42 黄胸鼠头骨量衡度 单位:mm

3)分类讨论 黄胸鼠(Rattus flavipectus)是Milne-Edwards(1871)订名的。Allen(1940)、Honacki(1982)均视其为独立种,并有2个亚种。而Ellerman和Morrison-Scott(1951)则认为它是黑家鼠(R.rattus)的一个亚种。国内学者认为,R.rattus与R.flavipectus在形态上有差异,在自然条件下未发现中间类型,在人工饲养下,二者也无杂种后代。所以,大多数学者均倾向于将其作为一个独立种。本书从此意见。
4)亚种分化 据Allen(1940)记述,黄胸鼠在国内有2个亚种,即指名亚种(R.f.flavipectus)和云南亚种(R.f.yunnanensis)。近来国内有些学者认为云南亚种与指名亚种没有显著差异,指出黄胸鼠在国内只有1个亚种即指名亚种,故有关其亚种分化还需进一步研究。本书暂从Allen(1940)等的意见。
河南省分布有1个亚种——指名亚种(R.f.flavipectus,Milne-Edwards1871),本亚种主要特征见种级记述。见于灵宝、新乡、济源、罗山、商丘、开封等地。
5)地理分布 黄胸鼠在河南省分布几遍全省各地(图4.73)。国内主要分布于长江以南诸省(区)。但在陕西、宁夏、新疆、甘肃等北方省(区)也有分布。国外尚无分布。
6)生态 黄胸鼠主要栖息于居民住宅、家庭院落等处。喜攀缘,多隐匿于房屋上层。常在屋顶缝隙、天花板、阁楼、房梁洞隙处营巢而居。室外柴草及杂物堆积处也可作为藏身之地,甚至火车、轮船上都有其踪迹。与人类伴居,也可分布于山林地带。1994年9月在济源太行山区调查时,海拔1 200 m处仍有捕获,捕获率为1.1%。另据调查(葛荫榕等,1987),黄胸鼠在豫东平原家栖鼠类中,可占到7.5%~14.4%。黄胸鼠多在夜间活动,但白天也时常出没。食性杂,但较偏于植物性食物。野外生活者,也爬树盗食鸟卵、雏鸟等。黄胸鼠的巢室构造比较简单,洞口直径4~5 cm。巢材包括碎纸、破布、棉絮、干草、植物叶等。全年均可繁殖。每年繁殖3~4胎,每胎产5~9仔。

图4.73 黄胸鼠的分布(▲)
7)经济意义 黄胸鼠是家栖主要害鼠之一。盗食粮食,污染各种食品,咬毁家具、衣物、电力器材等,甚至捕杀雏鸡、雏鸭。此外,还携带和传播流行性出血热等自然疫源性疾病,给人类正常生活和生命安全造成了极大危害和威胁,各地城乡均应予以大力灭杀。
4.3.4.4白腹鼠属Niviventer
河南省白腹鼠属分种检索表

(1)社鼠(Niviventer confucianus,Milne-Edwards 1871)
地方名:山老鼠。
英文名:Chinese white-bellied rat
1)鉴别特征 体型中等,身体修长,尾长超过体长,尾背腹两色,前后也为两色。耳较大,前折可达眼部。体背常具针毛,体腹面硫黄白色。
2)形态描述(图4.74)

图4.74 社鼠
①外形 体型中等大小,吻略短而尖。耳壳较大,前折可达眼部,尾长大于体长,尾部鳞环明显,夏毛具针毛,冬毛则针毛极少。前足掌部与后足跖部均裸露,跖垫与指(趾)垫发达,雌性具乳头4对,胸部2对,鼠鼷部2对。
②毛色 体背为棕褐色而略带铁锈红色调,在体背中部杂生黑褐色长毛,故颜色显暗。夏毛在体背、肩部等处有白色针毛,毛尖褐色。喉部常为纯白色。体腹面毛色多呈硫黄白色或乳白色,老年个体硫黄色调尤为浓重。四足背面白色,常具褐色斑,尾背腹两色,背方棕褐色,腹方白色。尾后部有一段较长的白色,约占尾长的1/3。
③头骨 头骨整体观狭长而低扁,吻部较长,鼻骨极度前伸,超出门齿唇面之外。颧弓比较细弱,不明显向外扩张。眶上嵴发达,颞嵴呈弧形向后延伸至鳞骨。枕骨近于垂直,外枕嵴明显。腭骨较短,后缘可及上臼齿列后缘连线。腭长小于或接近于颅全长之半。门齿孔较长而略宽,其后缘达上臼齿列前缘连线。听泡较小而扁平。顶间骨宽大,其宽约为长的2倍(图4.75)。
④牙齿 上门齿唇面橙黄色。第1上臼齿(M1)较大,约为第2、第3上臼齿(M2、M3)之和。其第一、第二横嵴正常,第三横嵴缺舌侧齿尖,共有8个齿尖。第2上臼齿(M2)第一横嵴仅留舌侧齿尖,第三横嵴缺舌侧齿尖,共有6个齿尖。第3上臼齿(M3)小,长度不及第1上臼齿(M1)之半,整个咀嚼面愈合成一个椭圆形齿环。
⑤量衡度 社鼠量衡度有关数据见表4.43、表4.44。

图4.75 社鼠的头骨
1.背面观;2.腹面观;3.侧面观
表4.43 社鼠外形量衡度

表4.44 社鼠头骨量衡度 单位:mm

续表4.44

3)分类讨论 自Milne-Edwards(1871)依据四川宝兴的该种标本订名为Mus confucianus(北社鼠)以来,社鼠的分类地位几经变动,主要表现为属级分类地位的变化,以及该种是否与Hodgson(1836)所描述的社鼠[Mus(Rattus) niviventer](模式产地尼泊尔加德满都)为同一种等问题上。Miller(1912)将鼠属Rattus与小鼠属Mus分开;Allen(1940)系统整理了学者们分别在Rattus、Mus及Epimys属下所订立的中国不同地区社鼠的种名和亚种名,将中国除台湾省所产的社鼠全部归并为Rattus confucianus,种下分4个亚种:指名亚种(R.c.confucianus)、山东亚种(R.c.sacer)、河北亚种(R.c.chliensisr)和海南亚种(R.c.lotipes)。Ellerman(1941,1947,1961)、Ellerman和Morrison-Scott(1951)认为Rattus confucianus与Mus(Rattus) niviventer为同物异名,将其作为后者的亚种。国内学者多从此意见。汪松等(1981)比较了文献记述及所见西藏与尼泊尔、印度接壤地区的社鼠标本,冯祚建等(1986)比较了Ellerman(1961)所列尼泊尔、锡金及缅甸等地区的R.niviventer与国内R.confucianus的标本的形态特征(包括枕鼻长短、听泡大小及其与枕骨长的比例等),也认为中国的社鼠与国外所产的应属同一物种,应列为R.niviventer的亚种。
另有一些学者对niviventer等物种的属级分类地位提出质疑。Misonne(1969)依据包括R.niviventer在内、过去被Ellerman(1949)归并成为刺毛鼠亚属(Maxomys)的各类在牙齿特征上与其他属各类存在的差异,将Maxomy提升为属;Musser(1979)则认为这个新属名仅对rajah组鼠类才是正确的(模式种为M.gartelsii)。Marshall(1976)将niviventer等与Maxomy属中其他分开,提出一个新亚属(Niviventer),并恢复confucianus的独立种的地位。Musser(1981)则进一步将其提升和确立为属,并给出了系统评述,他分析比较了Niviventer和Rattus在外形(如尾二色特征及尾尖毛束存在与否)、头骨特征(包括颧板、腭桥、翼蝶骨、中翼骨窝、翼窝、听泡、耳咽管和下颌骨形态、颧骨鳞骨根在颅骨的位置高低、听泡在鼎骨的附着状况等)、上下颌臼齿的形态结构和核型等方面的差异。同时,Musser也指出,关于这个新属以及该属中一些物种的界限尚有不同意见,如Niviventer niviventer与Niviventer confucianus的关系问题,尚待进一步的分类订正工作予以解决。Musser认为,目前尚无令人信服的证据表明分布于尼泊尔的种群与其更东部地区的为同一物种;在进一步分类订正之前,niviventer仅指由巴基斯坦东北部向东至锡金分布的类群,而confucianus则指分布于缅甸北部、泰国北部、中国大陆地区的类群。此后,国外学者多采用Niviventer confucianus对中国所产的社鼠命名(Walker,1983; Corbet and Hill,1992; Wilson and Reeder,1993)。Wilson和Reeder(1993)指出,Niviventer confucianus通常被归于Niviventer niviventer中,但现在的研究结果不支持这一划分意见。国内学者仅有黄文几等(1995)采纳了Marshall(1976)和Musser(1981)的意见。
4)亚种分化依据Allen(1940)的划分、汪松等(1981)的订正建议、冯祚建等(1986)的意见,中国的社鼠共分化出8个亚种。
①指名亚种(N.c.niviventer,Milne-Edewards 1871)本亚种的模式产地为四川宝兴。该亚种背毛赭黄色显著,腹毛黄白色。刺毛在冬夏季变化明显,夏季发达。尾长明显超过体长。尾长与体长的比例,依Allen(1940)的数据为125%~138%,平均为130%;依据汪松等(1981)的数据为101%~157%,平均为124%。足背大多为纯白色,少数为暗色。分布广,见于西南、华南、华中等地。
②河北亚种(N.c.chiliensis,Thomas 1917)该亚种的模式产地为河北省的清东陵。其背毛沙黄色或土黄色显著。刺在夏季发达,冬季细弱。尾较短,尾长与体长的比例平均为94.8%(88%~107.4%,Allen,1940),而汪松等(1981)的数据则为84%~125%,平均为103%。后足色较暗。分布于河北东北部至吉林西北部。
③山东地区(N.c.sacer,Thomas 1908)该亚种模式产地为山东烟台,其体背毛色和刺毛变化似河北亚种,但尾较长。尾长与体长的比例平均为127%(103%~151%);而汪松等(1981)的数据为120%(103%~135%)。Howell(1929)认为该亚种为上述两亚种间的过渡类型。分布于山东、山西、河南、陕西、甘肃等黄河流域地区。
④海南亚种(N.c.lotipes,Allen 1926)该亚种的模式产地海南那大。其体背具犯棕色调,腹毛黄白色。刺毛终年发达。脑颅隆起。尾长与体长的比例似指名亚种,平均为127%(109%~137%)(Allen,1940);汪松等(1981)的数据为126%(110% 140%)。分布于海南。
⑤缅甸亚种(N.c.mentosus,Thomas 1916)该亚种的模式产地为缅甸。其体背呈暗棕褐色,多数个体具胸斑,尾呈明显二色或具白色尾梢。尾较长,尾长与体长的比例平均为124.8%(104%~139%)(冯祚建等,1986)。后足背面具暗色斑。分布于西藏东南部和雅鲁藏布江的下游地区。在国外则主要分布于缅甸北部和泰国北部地区。
⑥玉树亚种(N.c.yushuensis,Wang et Zheng 1981)该亚种的模式产地为青海玉树。其体型较大,体长平均为152 mm(129~185 mm)。体背灰色色调显著,呈灰褐色或鼠灰色。刺毛仅见于夏季,且不如其他亚种者粗硬。尾之二色不甚明显,且无白色尾梢。尾长超过体长,其比例平均为115%(100%~136%)。该亚种的伴人能力强于其他亚种,栖息于住宅、仓库和村寨附近的农田中。分布于青海东南玉树地区、四川西北部及西藏东部的昌都地区北部。
⑦闹牛亚种(N.c.naaoniuensis,Chang et Zhao 1984)该亚种的模式产地为吉林西部洮安县闹牛山。其主要特征为尾在已知的各亚种中最短,尾长与体长的比例平均仅为87%(73%~89%)张子郁等(1984)。分布于吉林西部的闹牛山。
⑧台湾亚种(N.c.calturatus,Thomas1917)该亚种的模式产地为台湾阿里山。其体型较大。体色较大陆各亚种深暗,体背灰褐色或鼠灰色显著。尾二色明显,尾端白色。尾长显著超过体长。分布于台湾。也有学者将其作为独立的种(Thomas,1917; Ellerman,1941; Wilson and Reeder,1993)。
河南省分布有2个亚种。
指名亚种(N.c.confucianus),本亚种主要特征是体背毛色赭黄色浓重,足背中央暗褐色,仅尾端为白色。见于桐柏、商城、罗山等地。
山东亚种(N.c.sacer),本亚种主要特征是体背毛色沙黄色显著,足背正中央色淡,近白色。尾后段1/3~1/2部分为白色。见于灵宝、济源等地。
5)地理分布社鼠在河南省分布于豫西黄土台地、豫西北太行山地、豫西伏牛山地、豫南桐柏-大别山地等(图4.76)。国内除黑龙江、辽宁、新疆等省(区)外,各省(区)均有分布。国外分布于印度、缅甸、斯里兰卡、中南半岛等国家和地区。社鼠为东洋界常见种类之一,在亚洲东南部分布极为广泛。

图4.76 社鼠的分布
●山东亚种(N.c.sacer) ▲指名亚种(N.c.confucianus)
6)生态 社鼠多栖息于山地、丘陵地带的林区、灌丛及草丛等环境中,靠近丘陵的农田中也有发现。多营巢于林下灌丛、倒木及林缘石隙中,食性较杂,常以坚果如桐子、松子等为食,也吃一些野果、嫩叶及昆虫等。据1994年在罗山县董寨林场的调查,在农作物成熟收获季节,社鼠还迁入农田。社鼠主要营夜间活动,白天也时有见到。每年春末夏初为繁殖盛期。每胎产4~10仔,平均4~5仔。在桐柏山地捕获率可达10%。
7)经济意义 社鼠为林区优势鼠种之一,常对苗圃、树木更新和农业生产造成极大危害。在种群数量较大时,应采取有效措施,予以集中防治。
(2)安氏白腹鼠(Niviventer andersoni,Thomas1911)
地方名:山老鼠。
英文名:Anderson’swhite-bellied rat
1)鉴别特征 体型中等大小,形似社鼠但略大。体腹面毛纯白色,绝无硫黄或污黄色。尾长大于体长显著,腭长多在20 mm以上。第一上臼齿(M1)宽度多大于2 mm。
2)形态描述
①外形 体型中等,形似社鼠,但尾较长;耳大而薄,前折可遮住眼部。后足较长。雌性具乳头4对。
②毛色 体背毛色棕褐色或赭褐色,毛基深灰色,毛尖赭褐色,并杂生较多的黑色针毛,故体背毛色较深。眼部前后直至耳基部为灰褐色,体侧赭色较浓。尾基部的背方和腹方均为黑褐色,但其余部分腹方多为白色,尾的后段近1/2部分为白色。
③头骨 头骨整体观较社鼠者为大。鼻骨狭长,前端超出上门齿唇面,其后端略超过前颌骨上支的向后延伸部分。顶间骨较宽,眶上嵴较细弱。颞嵴呈弧形,向后延伸至顶骨外侧中部逐渐消失。颧弓较细弱。听泡小而扁平。门齿孔较长,其后缘超过上臼齿列前缘连线(图4.77)。

图4.77 安氏白腹鼠的头骨
1.背面观;2.腹面观;3.侧面观
④牙齿 第1上臼齿(M1)极大,宽度多在2 mm以上,其前外侧齿尖不发达。第一横嵴外侧齿尖几近消失。第三横嵴缺内侧齿尖。第2上臼齿(M2)的第一横嵴仅留内侧齿尖。第3上臼齿(M3)较小,内侧具3个齿尖,外侧具1个齿尖,上臼齿列较长。
⑤量衡度 安氏白腹鼠量衡度有关数据,见表4.45、表4.46。
表4.45 安氏白腹鼠外形量衡度
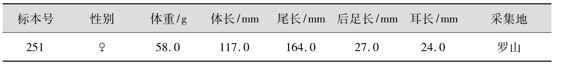
表4.46 安氏白腹鼠头骨量衡度 单位:mm

3)分类讨论 安氏白腹鼠是Thaomas(1911)依据采自四川峨眉山的标本所订立的,目前,多数学者倾向于将其作为独立的种(Honacki,1982;黄文几等,1995;王应祥,2003)。该种与社鼠N.confucianus在形态上很相似,但依前述的鉴别特征,二者仍可区分。罗泽珣等(1965)认为在同域分布环境中,该两种鼠的形态特征、睾丸长度的季节变化、年龄结构及生活习性等均有差异,故应作为两个独立的种。
4)亚种分化有关安氏白腹鼠的亚种尚待进一步探讨。
5)地理分布安氏白腹鼠在河南省分布于豫南大别山地(图4.78)。国内分布于陕西、台湾、云南、四川、西藏、甘肃等省(区)。国外分布于缅甸等国家。

图4.78 安氏白腹鼠的分布(▲)
6)生态安氏白腹鼠主要栖息于亚热带山地林区。常见于针阔叶混交林、阔叶林或近丘陵地区的农田、草地。食物以植物的茎、叶等绿色部分为主,也取食少量鲜果和昆虫等。
7)经济意义 安氏白腹鼠数量较少,为河南省稀有鼠类,故对农、林业的危害尚不足为害。
4.3.4.5 小泡鼠属Leopoldamys
小泡巨鼠(Leopoldamys edwardsi,Thomas 1882)
地方名:大山鼠、长尾巨鼠
英文名:Edward’s rat
(1)鉴别特征 体型粗大,为啮齿动物中体型较大者。成体体长接近300 mm。尾粗而长,尾长超过体长。耳大而薄,长可达38 mm,向前拉可遮住眼部。耳之背面生有棕褐色细绒毛。后足长56 mm,乳头4对。
(2)形态描述(图4.79)

图4.79 小泡巨鼠
1)外形 体型较大,为鼠科中的大型鼠类。尾粗而长,远超过体长。耳大而薄。口须刚硬而长。后足较长;乳头4对,包括胸部2对,鼠蹊部2对。
2)毛色 体背至头顶及尾基部为茶褐色,背部中央杂有黑色针毛,毛色较深,略呈黑褐色。体侧毛色略呈棕褐色。腹毛白色,背腹之间具明显的分界线。前后足的背面呈棕褐色。尾部背面为黑褐色,腹面为白色。
3)头骨 头骨粗壮而狭长,棱嵴发达,门齿孔后端达不到臼齿前缘的水平线。颅顶部略凸出;眶上嵴和颞嵴发达,向后外侧扩展直至顶间骨两侧角。腭骨后端接近第3臼齿水平线;眶下孔呈长三角形。鼻骨狭长,前端略超出门齿,前宽后窄;颧弓相对较细;听泡较小而低平(图4.80)。
4)牙齿 上门齿向后下方倾斜。门齿背面为棕黄色,侧面和腹面为牙白色。第1上臼齿(M1)较大,接近第2、第3上臼齿(M2、M3)之和。第1上臼齿(M1)第1横嵴外齿突不发达,中齿突最大,内齿突向下弯曲。第2横嵴3个齿突发育正常,内齿突仍向下弯曲,第3横嵴内外齿突消失,中齿突发达。第2上臼齿(M2)仅余一内侧齿突。第2、3横嵴与第1上臼齿(M1)第2、3横嵴相似。第3上臼齿(M3)第一横嵴亦仅有1内侧齿突,第2、3横嵴连接成环状。
5)量衡度 小泡巨鼠量衡度有关数据,见表4.47、表4.48。

图4.80 小泡巨鼠的头骨
1.背面观;2.腹面观;3.侧面观
表4.47 小泡巨鼠外形量衡度

表4.48 小泡巨鼠头骨量衡度 单位:mm

(3)分类讨论 种级分类地位无争议。Thomas(1822)将小泡巨鼠归于家鼠属Rattus,目前学者接受将其列为小泡鼠属(Leopoldamys)。
(4)亚种分化小泡巨鼠在国内已知有3个亚种:
指名亚种(L.e.edwardsi),由Thomas于1882年命名,其模式产地在福建西部山地。在中国大陆分布于广西、广东、安徽、福建、浙江、湖南等地。
四川亚种(L.e.gigas),由Satunin于1903年命名,其模式产地在四川。在中国大陆分布于陕西、贵州、云南、四川等地。
海南亚种(L.e.hainanensis),由Wu et Yu于1985年命名,模式产地在海南坝王岭、吊罗山。在中国大陆分布于海南等地。
由于标本数量较少,有关河南标本的亚种归属尚待进一步研究。
(5)地理分布 小泡巨鼠目前已知分布于豫西伏牛山区(图4.81)。国内见于陕西、甘肃、浙江、安徽、湖南、湖北、贵州、四川、云南、福建、广东、广西、海南等省(区)。国外分布于印度、印尼、老挝、马来西亚、缅甸、泰国和越南等国家。

图4.81 小泡巨鼠的分布(▲)
(6)生态小泡巨鼠主要栖息于中低山区的阔叶林、针阔叶混交林稀疏的林缘地带或高山密林的山谷中,但一般均靠近水源。也见于农林交错区的林缘灌丛或农田中。其巢窝多位于岩缝中;由于经常出入洞穴,洞口被磨得很光滑,常留有活动的足迹和粪便;洞外的草丛中常见有外出活动的跑道。春夏季活动范围较大,常到山区的田间去采食;并以灌丛、草丛等作为临时的隐蔽、休息场所。多在夜间活动,天黑以后即外出觅食。攀缘能力强,常爬到树上采食鲜果。主要食物是植物的茎、叶及幼嫩细根;出到农田取食玉米、甘薯、稻谷及各种野果。小泡巨鼠喜食蘑菇、竹笋、肉类等。在冬季,由于食物短缺,外出觅食活动减少,多在洞内休息。每年于夏、秋季繁殖两次,每胎2~4仔。
(7)经济意义小泡巨鼠是山区农林业害鼠,不仅危害各种野果,啃食树苗幼芽,给农林业造成危害,同时盗食竹笋、香菇、板栗等;春末夏初常进入农田,破坏农作物和蔬菜。但因个体较大,其毛皮可资利用。在河南省分布数量较少,无重大价值。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。